4º ESO Biología y Geología UD 6 Origen y evolución de la vida
1. Origen de la vida
Como podemos imaginar a lo largo de la historia las preguntas acerca de nuestro origen (y por tanto el de los seres vivos) han sido un tema de mucho interés al ser el ser humano un animal consciente de sí mismo y de su existencia limitada en el tiempo.
Todas las sociedades han desarrollado un corpus de creencias religiosas en las cuáles venía reflejado (entre otras cosas) la creación de los seres vivos y el hombre por una serie de seres superiores o dioses. Naturalmente estas mitologías eran diferentes en cada sociedad (aunque muchas veces estaban influidas por las mitologías de las sociedades con las que estaban en contacto y por las sociedades predecesoras).
Llamamos al conjunto de estas teorías religiosas comoteorías creacionistas o creacionismo. Todas ellas comparten el hecho de que la vida es otorgada o creada por un ser superior.
"La creación de Adán" de Miguel Ángel
En nuestra cultura particular (la judeocristiana) los mitos de la creación vienen recogidos en el libro del Génesis de la Biblia. Actualmente intenta venderse, sobre todo en USA, por parte de ciertos grupos religiosos cristianos esta idea revistiéndola de un vocabulario pseudocientífico con el nombre de “diseño inteligente”. Sin embargo sigue careciendo de toda base científica y se derrumba cuando se profundiza en el conocimiento de como funciona la vida y los procesos que llevan a cabo los seres vivos.
El estudio de las diferentes mitologías (griega, maya, india, japonesa, navajo, …) es sin duda apasionante pero escapa al alcance de este tema. Vamos a saltar por lo tanto al siglo XVII en Europa donde empieza a nacer la ciencia y con ella un intento de describir y comprender el mundo que va a buscar respuestas reales y objetivas.
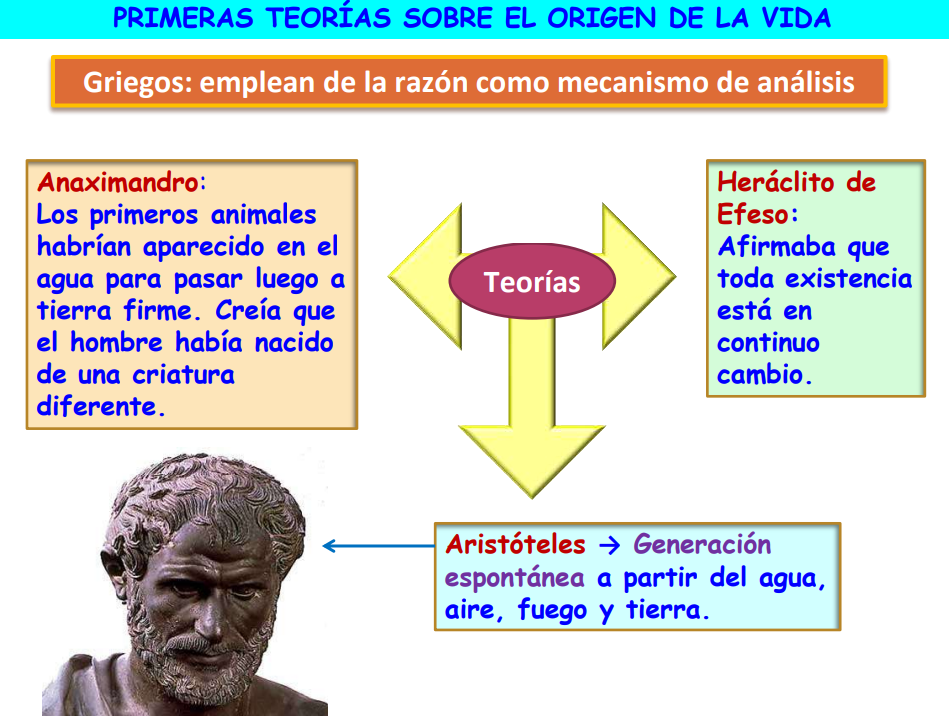
1.1. Teoría de la generación espontánea
Una de las teorías más antiguas sobre la vida es la teoría de la generación espontánea. Como su nombre indica esta teoría defendía que la vida, y por tanto los seres vivos, aparecían de la nada o a partir de la materia inerte.
Esta teoría proviene de la idea de Aristóteles de que la una fuerza vital o soplo divino (entelequia) animaba la materia inerte confiriéndole vida. El pensamiento filosófico de Aristóteles es de gran importancia en Europa durante toda la edad media y por eso esta teoría se acepta de forma mayoritaria.
El médico flamenco Van Helmont en 1667 incluso escribe una receta: “Si colocamos ropa interior sucia junto con trigo en un recipiente de boca ancha al cabo de 20 días el olor penetra en las cáscaras de trigo convirtiéndolo en ratones”.
Hoy en día esto nos parece absurdo e incluso por aquel entonces pocos aceptaban que fuera a aparecer un ciervo de repente de la nada pero la observación de la aparición de gusanos en la carne, de moho o de seres microscópicos en el agua de los charcos (cuando ya había microscopios para verlos) eran de difícil explicación en esa época.
1.1.1. Experimento de Redi
El médico italiano Francesco Redien 1668 demostró que los gusanos que aparecen en la carne descompuesta no aparecían de forma espontánea si no que eran larvas de moscas que procedían de los huevos que estas ponían en la carne muerta. Lo hizo con este sencillo experimento.
1.1.2. Experimento de Spallanzani
El experimento de Redi demostraba que los animales no aparecían por generación espontánea pero muchos científicos y pensadores pensaban que los microorganismos (recientemente descubiertos y estudiados gracias al desarrollo de los microscopios) sí que aparecían por generación espontánea.
Lázaro Spallanzani ideó un experimento para demostrar que los mismos eran transportados por el aire:
Se hervían dos frascos con caldo de carne para matar a los microorganismos: un abierto y otro cerrado. En el abierto aparecían microorganismos al cabo de pocos días y en el cerrado no aparecían nunca.
1.1.3. Experimento de Needham
Los experimentos de Redi y Spallanzani supusieron un fuerte golpe para la teoría de la generación espontánea pero científicos como Needhad sostenían que en el aire había una fuerza vital (esa entelequia de la que hablaba Aristóteles) y que al tapar los botes y calentarlos esta fuerza vital se eliminaba por el calor. Para probarlo hizo el siguiente experimento: hervía caldo de carne en un matraz para matar a los microorganismos y después lo tapaba con un corcho. Al cabo de unos días lo observaba al microscopio y encontraba seres vivos en el caldo. Esto probaba, según él, que estos seres vivos aparecían por la “fuerza vital” que había en el aire.
1.1.4. El experimento de Pasteur
La teoría de la generación espontánea queda definitivamente refutada y abandona con los magníficos y elegantes experimentos realizados por el químico francés Louis Pasteur.
Gracias a estos experimentos se postula la ley de la biogénesis que establece que todo ser vivo procede de otro ser vivo preexistente.
Al permanecer sus matraces abiertos en todo momento no se podía argumentar que se impedía a la “fuerza vital” pasar o que se la destruía por el calor. Los experimentos de Pasteur son tan estupendos que todavía hoy hay multitud de matraces abiertos de su época con el caldo de carne sin estropearse.
1.2. La teoría abiogénica de Oparin-Haldane
A principios del siglo XX, el bioquímico ruso Oparín y el biólogo escocés Haldane elaboraron, de forma independiente, una teoría abiogénica (proceso de aparición de la vida a partir de moléculas orgánicas simples) que trataba de explicar como había aparecido el primer ser vivo. Oparin tiene la primacía pues la publicó en 1924 y Haldane en 1927 pero la publicación de Oparín no llegó a occidente hasta los años 30.
En primer lugar había que explicar como se formaba materia orgánica a partir de materia inorgánica; esto requiere energía y compuestos reductores como el amoniaco o el metano que no se encuentran hoy en la Tierra en estado libre.
Según Oparín esto habría sido posible gracias a que en esa época la atmósfera terrestre no era oxidante como ahora sino reductora. La aparición del primer ser vivo habría tenido las siguientes etapas:
1)
Hace 4 000 Ma, los componentes de la atmósfera
reductora primitiva (metano, amoniaco, hidrógeno y vapor
de agua) reaccionaron gracias a la energía de las tormentas, la
radiación solar y las continuas erupciones volcánicas dando lugar a
moléculas orgánicas sencillas.
2)
Posteriormente, al enfriarse la Tierra, la lluvia arrastró estas
moléculas y formó los mares, que Oparín llamó “sopa
o caldo primordial”.
3)
Las moléculas orgánicas formadas se fueron uniendo hasta originar
otras más grandes. Algunas de ellas se aislaron del medio acuático
en el interior de estructuras denominadas coacervados.
4)
Algunos de estos coacervados desarrollaron en su interior moléculas
con capacidad para autorreplicarse (ácidos nucleicos). Fueron los
primeros organismo o progenotas,
origen de todos los demás.
1.2.1 El experimento de Miller
El científico norteamericano Miller desarrolló un experimento que replicase la composición gaseosa de la atmósfera primitiva y les aplicó descargas eléctricas de alto voltaje (equivalentes según él a los relámpagos) y obtuvo moléculas orgánicas sencillas como aminoácidos.
Hoy en día se cree que la atmósfera terrestre no era rica en amoniaco ni metano lo cuál haría que este experimento perdiese validez. Tampoco los polímeros se forman con facilidad en las condiciones descritas en su teoría.
Ha habido una reformulación de la misma trasladando el origen de la vida las fuentes hidrotermales o a capas finas de arcillas del litoral que actuarían como catalizadoras de las reacciones químicas.
1.3. Teoría de la panspermia
El científico Arrhenius atribuye el origen de la vida en la Tierra a la llegada de esporas (o similar) en el interior de meteoritos venidos del espacio. Esta teoría presenta muchos problemas desde la imposibilidad de encontrar ninguna prueba, pasando por cómo habían llegado esas esporas al interior de los meteoritos hasta la posibilidad de que sobrevivieran a la entrada en la atmósfera terrestre. Además tiene el inconveniente de que deberíamos preguntarnos cómo se origino esa vida en otro lugar.
Más interés tiene la panspermia molecular que afirma que hubo una llegada de moléculas orgánicas a la Tierra desde el espacio. Esto es muy posible dado que se han encontrado moléculas orgánicas en numerosos planetas, nebulosas, ...
1.4. Condiciones iniciales de la vida
Una de las cosas que parecen más claras hoy en día es que el primer ser vivo del que descienden todos los actuales se originó en las fumarolas submarinas o en fuentes hidrotermales. Tanto los restos fósiles más antiguos como la composición del citoplasma como la composición isotópica parece indicarlo.
1.5. Teorías actuales
“Existen numerosas teorías actuales que podemos agrupar en dos grandes corrientes:
a) la corriente del “metabolismo primero”; da prioridad al establecimiento de ciclos metabólicos cerrados de forma espontánea. Un ejemplo de esta corriente sería la teoría del hierro-sulfuro de Wächtersshäuser.
b) la corriente de los “genes primero”; da prioridad a la aparición de moléculas de ARN autocatalítico (ribozimas) que se verían englobadas en microsferas. Un ejemplo de esta teoría sería la teoría del mundo de ARN de Gilbert”
Ambas corrientes tienen numerosos inconvenientes hoy por hoy irresolubles. Existen numerosas teorías híbridas entre ambas y que intentan explicar diferentes aspectos como la teoría de la burbuja, la teoría de la arcilla, etc etc.
Debido a las ideas creacionistas y religiosas, en Europa siempre se había asumido que los seres vivos habían sido los mismos desde la creación de la Tierra (algo ayudado por la estimación de la edad de la Tierra en solo unos miles de años).
El gran botánico sueco Linneo (que ya estudiamos en 1º al ser el encargado de crear la nomenclatura binomial que se usa hasta hoy en día para dar el nombre científico a las especies) fue el primer científico de renombre que propugnó la teoría de que las especies habían sido creadas de forma separada e independiente y negó la posibilidad de un origen común de los seres vivos.
Es paradójico que la creación por parte de Linneo de la taxonomía y la agrupación de los seres vivos en géneros, familias, órdenes, …. fuera algo básico para el establecimiento de relaciones y parentescos entre los seres vivos y, por lo tanto, para las ideas y teorías evolucionistas posteriores.
2.1.1. El catastrofismo
A finales del siglo XVIII, Cuvier elaboró una teoría para intentar explicar uno de los fenómenos que más chocaba con la teoría fijista: ,cada vez se encontraban más fósiles de seres vivos que no se correspondían con seres vivos actuales (incluso algunos tan llamativos como los de los dinosaurios).
Según Cuvier se habían sucedido hechos catastróficos, el último de los cuáles habría sido el diluvio universal descrito en la Biblia, que habían extinguido a todos los seres vivos y posteriormente Dios volvía a crear nuevos seres vivos. Una de las pruebas que esgrimía del diluvio universal es la existencia de fósiles marinos en montañas y lugares muy lejanos del mar.
Estos grandes científicos encontraban pruebas y pistas que indicaban que los seres vivos cambiaban con el tiempo y estaban relacionados de forma más o menos cercana pero los profundos sentimientos religiosos de la época hacían muy difícil asumir la idea de una creación imperfecta por parte de Dios.
2.2. Pruebas de la evolución
Las teorías evolucionistas son teorías científicas modernas y, por lo tanto, deben basarse y apoyarse en pruebas. Vamos a estudiar las más importantes:
2.2.1 Pruebas anatómicas y morfológicas
Estas pruebas se consiguen mediante el estudio de la anatomía comparada entre los organismos vivos y también entre estos y los restos fósiles. De este estudio podemos destacar tres tipos de órganos que nos indican la evolución de las especies:
a) Órganos homólogos; son órganos que, aunque realizan distintas funciones, tienen el mismo origen evolutivo y comparten idéntico patrón estructural. Constituyen una prueba de laevolución divergente o radiación adaptativa, que consiste en la aparición de diversos cambios, de acuerdo con las diferentes formas de vida, a partir de un grupo antecesor.
b) Órganos análogos;son órganos que, aunque tienen diferente origen evolutivo, realizan la misma función. Constituyen una prueba de la evolución convergente, que consiste en cambios que han dado lugar a estructuras semejantes, en grupos no emparentados, a partir de distintos organismos antecesores.
c) Órganos vestigiales; son estructuras que tienden a desaparecer pues ya no resultan útiles a los seres vivos. Como consecuencia de su falta de utilidad se seleccionan aquellos individuos que dedican menos energía a las mismas y, por lo tanto, cada generación estarán menos desarrolladas. Constituyen una prueba de la evolución de los seres vivos a partir de seres vivos que vivían en unas condiciones ambientales diferentes o tenían unos hábitos de vida distintos.
2.2.2. Pruebas paleontológicas
Los fósiles revelan que han existido organismos diferentes a los actuales y, por lo tanto, que los seres vivos han cambiado a lo largo del tiempo.
Con el estudio de muchos paleóntologos y durante muchos años se ha conseguido encontrar fósiles de especies emparentadas que han permitido observar transformaciones progresivas. A esto se le denomina serie evolutiva y es una gran prueba de la evolución. Una de las mejor estudiadas y conocidas es la del caballo.
El estudio (que ya vimos en el tema 3) de los estratos (principio de superposición de los estratos) nos ayuda a ver la evolución temporal de fósiles más antiguos a más modernos (secuencia temporal).
Otra prueba de la evolución que aportan los fósiles son los eslabones evolutivos; son seres vivos con características que hoy poseen dos grupos distintos. Suponen la prueba de la relación y el paso de un grupo a otro.
Una última prueba paleontológicade la evolución son los llamados fósiles vivientes; estos son seres vivos que se han conservado sin apenas cambios durante muchísimo tiempo. Proporcionan más información que los restos fósiles y permiten estudiar en profundidad características primitivas.
2.2.3. Pruebas embriológicas
En 1866, Haeckel enunció la ley biogenética fundamental, según la cuál el desarrollo embrionario (ontogenia) es una recapitulación de toda la evolución de ese ser vivo (filogenia). Hoy en día se sabe que esta ley no es cierta pero sí es cierto que cuánto más similares son los embriones de dos especies más cercanas evolutivamente están.
2.2.4. Pruebas biogeográficas
Estas pruebas aparecen cuando estudiamos seres vivos que han quedado aislados e incomunicados por la aparición de barreras geográficas (mares, cordilleras, …). Cuanto mayor es el tiempo de aislamiento mayores diferencias aparecen entre esas especies cercanas.
2.2.5. Pruebas bioquímicas o moleculares
El estudio de la secuencia de aminoácidos en las proteínas o de nucleótidos en el ADN nos permite saber como de relacionadas están dos especies. Esto es especialmente útil cuando estudiamos moléculas que aparecen en muchas especies diferentes (por ejemplo, la hemoglobina en los vertebrados).
2.2.6. Pruebas coevolutivas
La coevolución es el proceso por el que dos especies se influyen mutuamente en su evolución. Por ejemplo:
a) Mutualismo; es una interacción entre dos especies en las que ambas obtienen un beneficio mutuo. La evolución de flores e insectos es un claro ejemplo de esta evolución. En muchos casos la evolución ha sido tan grande que solo un tipo de insecto poliniza un tipo de flor, y ese tipo de insecto solo va a ese tipo de flores.
b) Parasitismo;ocurre cuando una especie vive a expensas de la otra. Normalmente los parásitos son muy específicos y evolucionan diferenciándose del mismo tipo de parásito de otra especie. Incluso puede pasar en zonas distintas del cuerpo de la misma especie. Por ejemplo el piojo humano del pelo de la cabeza es distinto al del pelo corporal.
d) Domesticación;las especies que se domestican han sufrido enormes cambios como consecuencia de la selección artificial de aquellos caracteres más deseados por el ser humano. Un ejemplo muy efectivo es la enorme cantidad de razas de perros distintas creadas a partir de un antepasado común.
Hay fotos con más perros pero está es más cuqui^^
2.3. Teorías evolucionistas
Una cosa importante que tenemos que tener en cuenta es que la evolución es un hecho, como la gravedad, que ha sido demostrado mediante miles de experimentos. Las teorías evolucionistas no intentan demostrar que la evolución existe, igual que Newton o Einstein no intentaban demostrar que la gravedad existía; lo que intentan es explicar y averiguar cómo y por qué causas ocurre la evolución.
Vamos a ir estudiando las principales teorías por orden cronológico:
2.3.1. El lamarckismo
Esta teoría toma nombre de Jean Baptiste de Monet, caballero de Lamarck (1744-1829). Fue el primero en elaborar una teoría evolucionista razonada y se enfrentó a los defensores del fijismo encabezados por Cuvier.
Retrato de Lamarck
Al igual que Cuvier, Lamarck estudió los fósiles y observó que algunos mostraban aspectos intermedios entre los más antiguos y los actuales (como vimos en las pruebas paleontológicas). Esto le hizo pensar, acertadamente, que unos procedían de los otros.
La teoría de Lamarck es una teoría transformista que afirma que los seres vivos desarrollan órganos o estructuras y, por lo tanto, son capaces de transformarse.
Se apoya en tres hipótesis fundamentales:
a) Tendencia innata a la complejidad;según esta hipótesis, los seres vivos tendrían un “impulso interno” hacia la perfección y la complejidad (esta idea está muy vinculada con las ideas religiosas y el deseo de acercarse a Dios, que es la perfección). Este impulso sería el “motor” de la evolución y lo que provocaría la aparición de nuevos órganos y estructuras en los seres vivos para adaptarse a los cambios del medio ambiente. Esta tendencia se resume en su frase: “la necesidad crea el órgano”. Por eso a la teoría lamarckista también se la conoce como teoría de los caracteres adquiridos.
La evolución humana lineal sería un ejemplo de esa tendencia a la "perfección"
b) Hipótesis del uso y desuso; según esta hipótesis los órganos más útiles para el ser vivo se desarrollan y se potencian y los que no usa se atrofian y acaban desapareciendo.
c) Hipótesis de la transmisión de los caracteres adquiridos; según Lamarck todos los cambios (mejoras y atrofias) que un ser vivo desarrollaba durante su vida se los transmitía a sus descendientes. Gracias a esto, con el paso del tiempo y de muchas generaciones, los pequeños cambios del ser vivo se iban amplificando y convirtiendo en grandes cambios que explicaban la gran diversidad de formas de vida que hay en la actualidad.
Aciertos de la teoría de Lamarck
a) Admite un proceso evolutivo y propone un mecanismo para explicarlo.
b) Reconoce la relación entre las estructuras anatómicas y su función (Anatomía comparada).
Errores de la teoría de Lamarck
a) Afirmó que los seres vivos tienen la necesidad interna de perfeccionarse y hacerse más complejos (prejuicios religiosos).
b) Defendió la transmisión de caracteres adquiridos durante la vida del ser vivo (total desconocimiento en su época de la genética, el ADN, los gametos, etc etc)
Leer con atención. Es un ejemplo de sencillo experimento para refutar una teoría.
Últimos descubrimientos: la epigenética
En los últimos años se ha descubierto que, aunque la secuencia de bases del ADN no cambie, las circunstancias que afectan a un ser vivo pueden modificar su ADN inactivando o activando genes mediante metilaciones. Incluso puede ocurrir en las células germinales y, por lo tanto, transmitirse a los descendientes.
Esto podría sugerir una cierta transmisión de caracteres adquiridos aunque todavía es pronto para saber su influencia en los procesos evolutivos a medio/largo plazo dado que estas alteraciones pueden deshacerse con facilidad.
Esta teoría fue elaborada de forma independiente por Darwin y Wallace; Darwin la desarrolló primero pero no se atrevió a publicarla y Wallace le enseñó su teoría elaborada en la India con posterioridad. Cuando Darwin le enseñó a Wallace sus manuscritos éste le reconoció su primacía en un gesto que le honra. Finalmente Darwin, animado por el descubrimiento de Wallace y temiendo que alguien más se le adelantara publicó su teoría en uno de los libros científicos más importantes de la Historia: "El origen de las especies por selección natural".
Darwin
Wallace
Podéis echar un vistazo a sus entradas en la wikipedia para saber más de ambos naturalistas británicos.La teoría de Darwin se basa en los siguientes principios básicos:
a) Variabilidad intraespecífica; los individuos que conforman cualquier población de seres vivos presentan diferencias anatómicas, fisiológicas o de comportamiento.
Los individuos son diferentes
b) Supervivencia de los mas aptos; Siempre se producen más individuos en cada generación de los que pueden sobrevivir hasta reproducirse. Esto es debido a que los recursos son limitados (alimento, espacio, ...). Esto significa que una parte importante de los individuos mueren sin llegar a tener descendencia. Debido a esto habrá una competencia entre ellos por el espacio, el alimento y la reproducción.
c) Selección natural; aquellos individuos cuyas diferencias, o variaciones, les faciliten la supervivencia y la reproducción se verán favorecidos mientras que aquellos cuyas diferencias no les faciliten, o les dificulten, la supervivencia y la reproducción se verán perjudicados.
Las polillas oscuras en bosques contaminados son menos comidas por pájaros que las claras
d) Evolución de la especie; como los individuos con las diferencias más favorables sobreviven en mayor número, se van a reproducir en mucho más número y, por lo tanto, la siguiente generación se parecerá más a ellos que a los que no han podido reproducirse y han muerto. Esto se debe a que los descendientes se parecen a sus progenitores. Con el paso de mucho tiempo y muchas generaciones de individuos los cambios son cada vez más notables y pueden llegar a dar lugar a nuevas especies.
Aciertos de la teoría de Darwin
a) Darwin se acerca mucho más a lo que sabemos de la evolución que Lamarck.
Hacer dibujo bonito con explicaciones
b) Introduce los conceptos de variabilidad intraespecífica, competencia por los recursos y selección natural. Estos son motores fundamentales en la evolución y todavía sustentan las teorías evolutivas modernas.
Errores de la teoría de Darwin
a) Darwin no sabe explicar por qué existe la variabilidad intraespecífica y los individuos son diferentes. Algo normal dado que no se conocía el ADN ni nada sobre genética.
b) Tampoco era capaz de explicar como los caracteres pasaban de padres a hijos. El redescubrimiento de las leyes de Mendel y el nacimiento de la genética serán fundamentales para el estudio de la evolución.
c) Finalmente Darwin pensaba que los individuos eran capaces de adaptarse y cambiar (era transformista como Lamarck). Hoy en día sabemos que la que se adapta y cambia es la población y la especie a lo largo del tiempo (los individuos se quedan como nacen). Como ya vimos en la última clase, la epigenética ha venido muy recientemente a mostrar que sí existe un cierto transformismo heredable.
2.3.3. El neodarwinismo o teoría sintética de la evolución
Dos de los principales problemas de la teoría de Darwin era ignorar el origen de la variabilidad intraespecífica y como los caracteres de los progenitores pasan a sus descendientes.
Esta teoría coge la teoría darwinista y la une con todo lo aprendido sobre la reproducción sexual (y como consigue descendientes con combinaciones de caracteres diferentes a los progenitores), la recombinación genética, las mutaciones y las leyes de Mendel.
El neodarwinismo considera que la selección natural no cambia a los individuos sino a las poblaciones en su conjunto que ven afectada la frecuencia en que se encuentran unos individuos u otros (frecuencias génicas). Al ir acumulándose muchos cambios con el paso de mucho tiempo, las especies evolucionan y pueden llegar a dar lugar a nuevas especies.
2.3.4. El neutralismo
Kimura sostiene que la gran mayoría de mutaciones originan variaciones de genes que no son ni positivas ni negativas evitando que actúe sobre ellos la selección natural.”
Si esos individuos con genes diferentes quedan aislados del resto por algún motivo podría originarse una nueva especie. Esto haría que la evolución fuera un proceso muy regular. Hay que tener en cuenta que Kimura habla sobre todo de evolución molecular, no de individuo.
Los paleontólogos Eldredge y Jay Gould han propuesto esta teoría que afirma que, en muchas ocasiones, las nuevas especies surgen muy deprisa.
Obviamente, como son paleontólogos, se basa en el estudio de las series fósiles que muestran grandes periodos de estabilidad con periodos en los que aparecen grupos de fósiles que no parecen guardar relación directa con formas previas o con formas de transición entre un grupo y otro.
Achacan estos grandes cambios a macromutaciones que afectan a genes que regulan otros muchos genes unido a un ambiente con grandes nichos y espacios colonizables debido a grandes catástrofes o cambios climáticos.
Ponen como ejemplo de su teoría la aparición de los grandes grupos de seres vivos actuales en el Cámbrico o la aparición de los trilobites.
Esta teoría es compatible con la teoría neodarwinista y afecta solo a los ritmos, saltos y velocidades de la macroevolución.
2.3.6. Teoría Evo-devo (Evolutionary Developmental Biology)
Esta teoría se basa en el descubrimiento de los genes Hox que regulan la expresión de grupos de genes y de la organización de diferentes regiones del cuerpo durante el desarrollo embrionario.
Esto conlleva que mutaciones en grupos muy pequeños de genes serían responsables de grandes cambios corporales que darían lugar a nuevos grupos de organismos.
Un ejemplo sería el grupo de genes que controla el número, longitud y separación de los elementos esqueléticos tanto de las aletas de los peces como de las extremidades de los tetrápodos. Se piensa que de esta forma pudieron formarse los distintos tipos de extremidades que vemos en los vertebrados.
2.3.7. Teoría Eco-evo-devo (Ecological Evolutionary Developmental Biology)
Esta teoría supone una ampliación de la teoría Evo-devo al incorporar los factores ambientales como parte de los factores causantes del cambio evolutivo.
La epigenética estudia como factores ambientales (temperatura, fotoperiodo, dieta, ...) puede producir fenotipos característicos mediante una alteración en la expresión génica. Estudios recientes sugieren una cierta heredabilidad de esos cambios producidos por la epigenética lo que conllevaría que se convirtieran en actores evolutivos relevantes.
2.3.8. L.U.C.A. (Last Universal Common Ancester)
3. La especiación
Unaespecieestá constituida por un conjunto de individuos con un aspecto morfológico semejante, con las mismas características anatómicas y fisiológicas, y que se pueden reproducir entre sí y tener descendencia fértil.
La microevolución es el proceso por el que una especie se diferencia en especies muy semejantes que conforman un mismo género.
La macroevolución es el proceso por el que aparecen los grandes grupos de seres vivos mediante cambios muy grandes. Los gradualistas afirman que estos grandes cambios ocurren por la acumulación de pequeños tiempos en largos periodos de tiempo y los puntualistas que son bastante bruscos y no por la acumulación de pequeños cambios graduales.
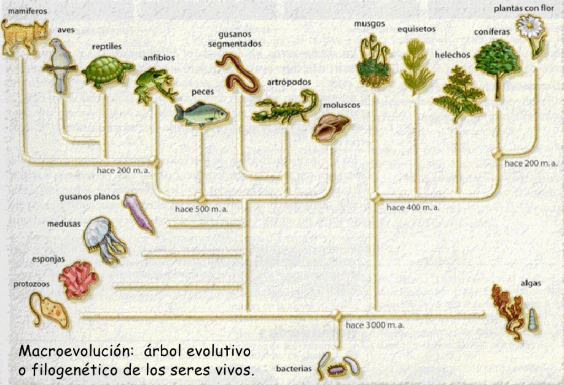
La filogeniaes el estudio de las relaciones evolutivas de los seres vivos. Normalmente se representan mediante un esquema simplificado que muestra las relaciones de parentesco mediante ramificaciones sucesivas (árbol filogenético).
La biodiversidad comprende la amplia variedad de seres vivos que existen en la Tierra.
3.1. Producción de cambios evolutivos en las poblaciones
Ya hemos estudiado que en las poblaciones existe una diversidad intraespecífica que hace que no todos los individuos sean iguales.
Está claro que los seres humanos no son idénticos. En todas las especies pasa lo mismo solo que nosotros no lo notamos tanto (ellos sí).
Como consecuencia de la selección natural uno de estos caracteres puede verse más favorecido que otros cuando hay algún cambio en el ambiente.
Cuando los bosques no estaban contaminados las polillas claras se camuflaban mejor y eran menos comidas por los pájaros. Por eso había muchas más.
Con la revolución industrial los árboles estaban sucios de hollín y las polillas negras se veían menos así que eran menos comidas y pasaron a ser mucho más abundantes.
Cuando se instauró la ley del aire limpio en Inglaterra los árboles (y las polillas) volvieron a su situación inicial. En el hilo de twitter os enseñan un magnífico experimento que prueba está situación:
Mirad que experimento más chulo que demuestra que la evolución no es cosa de dinosaurios, de fósiles ni de periodos de miles de años, sino en este caso, menos de un año. ¡Y no son bacterias! ¡Ni moscas! Es un vertebrado... Lo explico un poco mejor#scicomm#biodiversidadpic.twitter.com/WO7XJxaCkG
Este primer paso es importante pero no basta para formar una nueva especie pues los individuos todavía pueden reproducirse entre ellos y producirán individuos intermedios. Por eso las polillas blancas y negras siguieron siendo de la misma especie.
3.2. Aislamiento genético
Para que aparezca una nueva especie es imprescindible que los miembros de la nueva población no puedan reproducirse con los miembros de la población original. Cuando pasa esto deja de haber intercambio genético y mezcla entre las dos poblaciones. Deben aparecer unas barreras que impidan la reproducción y que pueden ser:
a) Barreras geográficas;impiden el contacto físico entre las poblaciones y, por tanto, la reproducción entre sus individuos. Ejemplos típicos son el aislamiento en islas o lagos, la separación por cordilleras o desiertos, etc.” En el hilo de twitter anterior serían las vallas que levantan los científicos.
b) Barreras sexuales; pueden ser debidas a diferencias anatómicas que impidan el apareamiento o a una falta de sincronía en los periodos fértiles. En cualquiera de estos casos, la reproducción no es posible aunque las poblaciones vivan en el mismo lugar.
c) Barreras fisiológicas; los gametos no son compatibles y aunque los individuos intenten reproducirse entre ellos no ocasionan descendencia.
d) Barreras cromosómicas; se producen cambios en el número y estructura de los cromosomas que hace imposible su reproducción. Es muy frecuente en las plantas y también se ha observado en algunos animales como el género Rana.
e) Barreras mecánicas; se producen cambios físicos en los aparatos reproductores que impiden la reproducción entre los individuos de las dos poblaciones.
Los órganos copuladores de estos caballitos del diablo son distintos y solo compatibles con su especie
Si los perros fueran animales silvestres sería imposible que pudieran reproducirse entre poblaciones tan distintas
f) Barreras etológicas; consiste en la aparición de nuevos comportamientos en ciertos individuos que producen rechazo a otros (normalmente cuando el macho intenta cortejar a la hembra). Cambios en el canto, cortejo, coloración, etc, son los ejemplos más típicos.
4. La aparición de la especie humana
4.1. Evolución de los primates
El Homo sapiens pertenece al orden de los primates, que apareció hace más de 60 Ma. y presenta una serie de adaptaciones comunes como resultado de su adaptación al medio arbóreo:
a)Visión estereoscópica con los ojos dirigidos hacia delante para moverse de rama en rama.
b) Antebrazo supinador con una estructura que les permite trepar.
c) Dedo pulgar oponible en las cuatro extremidades (la especie humana ha evolucionado hacia que el de los pies deje de estarlo) que les permite agarrar objetos.
d) Uñas planas en lugar de garras que facilitan la manipulación de objetos.
Dentro de los primates distinguimos dos grandes grupos:
4.1.1.Suborden estrepsirrinos; son capaces de sintetizar la vit. C, su olfato es mejor que la vista (muchos de ellos son nocturnos) y tienen un mayor número de dientes. Su morro es húmedo. Los más conocidos son los lemures que viven en Madagascar.
4.1.2. Suborden haplorrinos; no son capaces de sintetizar la vit. C y, por tanto, necesitan una dieta rica en fruta. Menor número de dientes y morro/nariz seco. Distinguimos dos grandes grupos separados por su distribución geográfica:
a) Infraorden Platirrinos o monos del nuevo mundo (americanos); tienen hocicos achatados con los orificios nasales hacia los lados, son especies de tamaño muy pequeño y conservan una cola larga.
b) Infraorden Catirrinos o monos del viejo mundo (africanos); tienen los orificios nasales hacia abajo separados por un tabique nasal, han perdido la cola o no es prensil. Estos primates pueden dividirse en dos grandes grupos:
· Superfamilia Cercopthidos; tienen cola (aunque no es prensil). Los más conocidos son los macacos de Japón o Gibraltar, los colobos y los babuinos.
Los macacos de Japón son capaces de sobrevivir al duro invierno bañándose en aguas termales
En Gibraltar encontramos a los únicos representantes europeos. Se han hecho muy confiados y suelen robar comida a las personas desprevenidas.
Los babuinos viven en grandes familias y sobreviven gracias a su inteligencia y trabajo en equipo
· Superfamilia Hominoidea; no tienen cola. Dentro de esta superfamilia distinguimos a la familia de los Hylobatidae, que comprende a los gibones, y a la familia Hominidae, que comprende a los homínidos, dentro de los cuáles está el hombre.
Gibón
4.3. Evolución de los homínidos
Dentro de la familia de los homínidos encontramos tres grupos:
4.3.1. Subfamilia Ponginae; actualmente solo tiene un representante vivo: el orangután (Pongo pygmaeus). Vive en las selvas tropicales cerradas de Java y Borneo. Tienen un marcado dimorfismo sexual: los machos pesan casi el triple que las hembras y tienen una cresta muy característica en el cráneo. El macho suele controlar un territorio en el que viven las hembras y sus crías.
Orangután macho
Orangután hembra con su cría
4.3.2. Subfamilia Gorillinae; actualmente solo hay dos especies de gorilas. Viven en selvas tropicales tanto de montaña como de tierras bajas. Tienen un marcado dimorfismo sexual siendo los machos entre el doble y el triple de pesados. Su gran peso les hace caminar apoyando los nudillos. Viven en familias con un solo macho dominante que controla un harén de hembras, sus crías y machos jóvenes inmaduros (cuando maduran son expulsados por el macho dominante y deben vivir solos hasta que puedan constituir su propio grupo familiar).
gorila hembra cuidando a su cría
gorila macho
Los gorilas de montaña fueron muy popularizados por la película “Gorilas en la niebla” que se realizó sobre el trabajo de la bióloga Diane Fossey asesinada por cazadores furtivos en 1985.
Podéis ver la película en Movistar+ y Filmin si tenéis alguno de los dos
4.3.3. Subfamilia Homininae; actualmente comprende a dos grupos:
a) género Pan; son los chimpancés, las especies más cercanas al hombre. Hay dos especies:
· El chimpancé común (Pan troglodytes); capaz de vivir en muchos hábitats distintos. Su alimentación básica es la fruta pero la complementan con pequeños animales que cazan en grupo, insectos, …. Presentan dimorfismo sexual pero mucho menor que los anteriores (30-40 Kg hembras y 40-60 Kg los machos). Sus piernas son cortas y se desplazan apoyando los nudillos como los gorilas. Su estructura social es compleja y forman grupos familiares variables en los que puede haber varios machos reproductores. Los machos cazan en grupo, defienden el territorio y se agrupan en grandes grupos (de hasta 150 individuos) o se disgregan en grupos más pequeños dependiendo de la necesidad."
· El bonono (Pan paniscus); vive en selvas tropicales de África central. A diferencia del chimpancé común cuya sociedad es patriarcal, los bononos tienen una sociedad matriarcal. Los machos del grupo pueden no estar emparentados y cooperan en la búsqueda del alimento. Todos pueden reproducirse con las hembras y suelen utilizar las relaciones sexuales frecuentes para aliviar tensiones sociales y evitar la competencia. Las hembras cooperan muy intensamente y establecen relaciones de amistad para cuidar a las crías y buscar alimento.
Si hablábamos del gran trabajo de la primatóloga Diane Fossey con los gorilas, los chimpancés han sido muy estudiados y popularizados por otra gran primatóloga: la doctora Jane Goodall. También se ha hecho varias películas sobre su trabajo:
Podéis verla en Filmin
Podéis verla en Netflix y Disney+
b) género Homo;actualmente solo comprende a una especie, la nuestra, Homo sapiens sapiens. Originalmente habitaba las sabanas africanas pero se ha extendido a todos los hábitats gracias a su tecnología. Su dieta es variada aunque la carne es mucho más importante que para los otros grupos. Tienen un dimorfismo sexual significativo siendo los machos de alrededor de 80Kg y las hembras de alrededor de 60Kg. Sus caninos son muy pequeños y no sobresalen. Sus piernas son más largas que los brazos, su cadera muy estrecha y carecen de pulgar oponible en los piés lo que explica su avanzado bipedismo. Su estructura social es muy compleja y su conducta sexual, relativamente parecida a la de los bononos, también.
4.4. Evolución de los homínidos no humanos
Decíamos en la clase anterior que chimpancés y bononos son las especies más cercanas al hombre de las que viven actualmente. Compartimos un ancestro común hace alrededor de 6 millones de años. Vamos a empezar a estudiar las ramas evolutivas que surgen a partir de entonces y acabarían dando lugar a los humanos modernos.
4.4.1. Ardiptihecus ramidus (4,4 m.a.)
Es el eslabón conocido más antiguo en nuestra historia evolutiva. Fue descubierto en 1992 por White en Etiopía.
Muestra características muy primitivas próximas a los ancestros del chimpancé como el esmalte fino en sus dientes, grandes caninos, … Sin embargo el desgaste de los dientes y una posible marcha bípeda (no segura) los aleja de ellos.
Este gran género comprende a especies anteriores o contemporáneas a los primeros representantes del género Homo.
Se caracterizan por tener todavía un cerebro pequeño, una dentición bastante potente y ser totalmente bípedos (algo fundamental en la evolución humana). Los más importantes son:
a) A. anamensis (4,2-3,9 Ma.); se han encontrado sus fósiles en Kenia y Etiopía. Su esmalte dental es grueso de lo que se deduce que comía alimentos duros. Vivía en ambientes forestales pero más abiertos que sus antepasados. Presentan un marcado dimorfismo sexual y aunque su mandíbula en forma de U es similar a la de los simios sus piernas muestran que ya era bípedo.
b) A. afarensis (3,9-3 Ma.); se han encontrado sus fósiles en el este de África (Etiopía, Tanzania y Kenia). Hoy en día se acepta que es uno de los ancestros directos del género Homo.
Su paladar tiene una forma intermedia entre U (simios) y V (hombre). Sus caninos eran pequeños y proyectados hacia delante (existe diastema) y su dentición a punta a una alimentación básicamente frutívora y herbívora.
Su cráneo es similar al de los chimpancés (400-550 cm3) y su cara presenta un fuerte prognatismo.
Su pelvis y sus piernas muestran que eran totalmente bípedos aunque sus brazos todavía eran más largos que sus piernas.
Marcado dimorfismo sexual con machos mucho más corpulentos que las hembras.
A finales de 1974Johansonencontró un esqueleto muy completo del, por entonces, homínido más antiguo encontrado. Lo llamó Lucy porque estaba sonando la canciónLucy in the sky with diamonsde los Beatles. Su descubrimiento y su estudio fueron uno de los principales hitos en el estudio de la evolución humana.
c) A. africanus (3-2 m.a.); se han encontrado sus fósiles en Sudáfrica. Presenta incisivos pequeños y caninos pequeños que no se proyectan hacia delante. Desaparece la diastema y sus molares y premolares tienen superficies planas.
Tienen un cráneo de 480-520 cm3 y su caja craneal es más alta y redondeada que la de los chimpancés. Su cara es más corta y menos prognata al tener dientes más pequeños.
Presentan una estatura de 1,50 m. Un peso entre 35-65 Kg (fuerte dimorfismo sexual). Y locomoción totalmente bípeda.
De acuerdo con el estudio de sus fósiles sería el tronco común del que surgen dos grandes ramas evolutivas: una hacia el géneroParantropusy otra hacia el géneroHomo.
Existen otras especies importantes como A. bahrelghazali y A. garhi pero no parecen estar directamente relacionadas con la línea evolutiva hacia el género Homo.
4.4.3. Unos primos lejanos: el género Paranthropus
Este género deriva, como el género Homo, de los Australopithecus. Vivieron en ambientes áridos y semiáridos alimentándose de vegetales duros gracias a un potente aparato masticador con el que trituraban raíces y semillas.
Su capacidad craneal es de alrededor de 520 cm3 y tenían una cara muy reforzada con enormes molares y grandes músculos masticadores. Los machos tenían una cresta sagital similar a la de los gorilas.
Convivieron con varias especies del géneroHomoy tuvieron bastante éxito evolutivo extendiéndose al sur de África pero, finalmente, se extinguieron sin dejar descendencia. Las especies más destacadas sonP. aethiopicus(2,6-2,2 m.a.),P. robustus(2-1,2 m.a.) y, sobre todo,P. boisei(2-1 m.a.).
4.5. Evolución del género Homo
Todo el proceso que conduce a la aparición de los humanos modernos se conoce con el nombre de hominización y los estudiaremos con detalle en el último punto del tema.
4.5.1. Homo habilis (el primer hombre) (2,4-1,8 m.a.)
Corporalmente no se diferencian mucho de los australopithecus pero su capacidad craneal es muy superior: 500-750 cm3. Esto le habría permitido tener mayores capacidades cognitivas y una mayor complejidad social.
Se le atribuye el desarrollo de las primeras industrias líticas que le permitieron incorporar muchos alimentos (sobre todo carne y huesos) a su dieta. Probablemente eran carroñeros oportunistas y gracias a estas herramientas de piedra podían aprovechar al máximo las carroñas raspando los huesos y extrayendo el tuétano de los mismos. Este gran aumento de proteínas y calorías permitiría su mayor capacidad cerebral.
Todavía no controlaba el fuego y carecía de un lenguaje articulado que le permitiera transmitir ideas complejas.
Sus dedos siguen siendo curvos lo que probablemente indica que todavía subían a los árboles. Sus caderas se han estrechado mucho para desplazarse mejor de forma bípeda. Esto hace que las crías tengan que nacer de forma prematura y requieran muchos más cuidados y durante mucho más tiempo que en la mayoría de los animales.
Su hábitat ya no es forestal y aparecen habitando espacios mucho más abiertos tipo sabana.
4.5.2. Homo rudolfensis (1,9-1,6 m.a.)
Presenta una mayor capacidad craneal que H. habilis. Un esqueleto facial más grande y plano y una mandíbula y dentición más potente.
4.5.3. Homo ergaster (1,8-1,4 m.a.)
Tiene rasgos humanos y complexión robusta. Son un gran paso desde los H. habilis y están mucho más cerca de los humanos modernos. Sus piernas ya son más largas que los brazos como en los humanos modernos.
Su capacidad craneal es mayor (800-900 cm3). Sus cráneos son alargados, bajos y con la base ancha; con arcos superciliares bien desarrollado separado del hueso frontal por un surco bien marcado, son menos prognatos y sus huesos nasales sobresalen del resto de la cara.
Se le atribuye la invención de las hachas bifaces (con dos caras). Achelense.
El esqueleto más completo encontrado de esta especie es el niño de Turkana (10-11 años de edad) con un tamaño y estructura corporal similar al nuestro.
Los ergaster ya son grandes cazadores y depredan incluso megafauna como los elefantes adulto. Esto supone un incremento enorme en la alimentación y en la disponibilidad de energía.
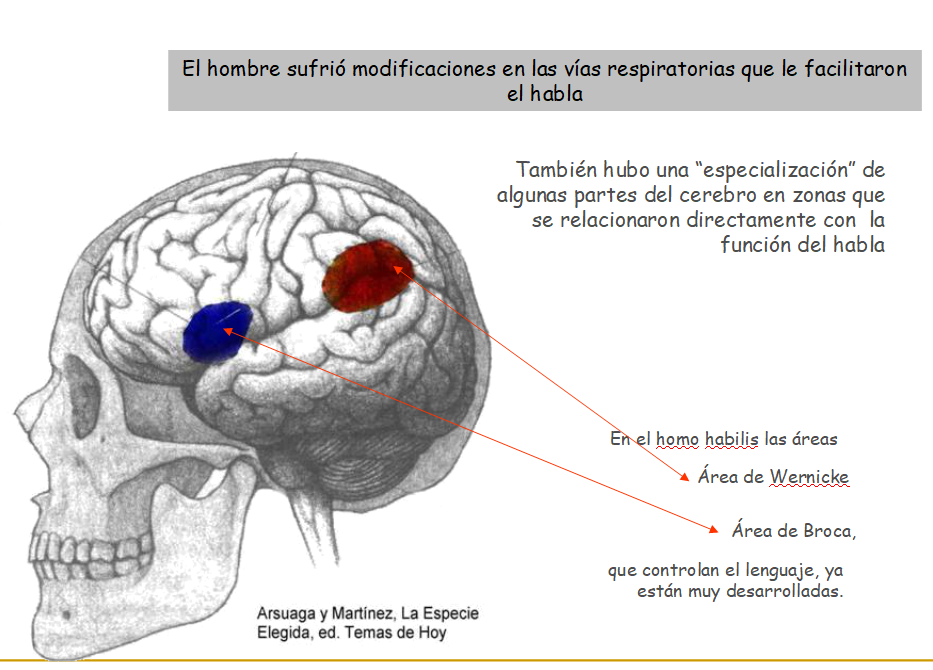
Existen dudas sobre si podían hablar. Su área de Broca (parte del cerebro dedicada al lenguaje) está bien desarrollada pero la estructura de su cuello hace que muchos investigadores crean que no tenía hueco para articular palabras. Su desarrollo infantil es más largo (como en los humanos modernos) lo que permitiría un mayor tiempo de aprendizaje. También permite que los lazos sociales y familiares se hagan más intensos.
Es la primera especie que sale de África y coloniza Europa, Asia, América y Oceanía. En Asia está especie evolucionará originando el Homo erectus y en Europa originando el Homo antecessor.
4.5.4. Homo erectus (1 m.a. -100.000 a.)
Esqueleto muy similar al nuestro. Cráneo más macizo, frente más pequeña y arcos supraciliares prominentes.
Son contemporáneos con los neardentales y los sapiens. H. erectus, H. neanderthalensis y Homo sapiens evolucionan en tres regiones distintas aisladas geográficamente (Asia, Europa y África).
Controlaban y utilizaban el fuego.
Extendidos por toda Asia y Oceanía. En las diferentes islas y regiones va a originar subespecies y variedades adaptadas a dichos medios cuando quedaban aisladas por subidas en el nivel de océano.
4.5.5. Homo antecessor (800.000 a. - 500.000 a. )
Esta especie fue descubierta por un equipo bajo la dirección del paleontólogo español Arsuaga en la Gran Dolina en Atapuerca (Burgos). Posteriormente se han encontrado fósiles en Francia e Inglaterra que podrían coincidir aunque no hay evidencias concluyentes.
Eran individuos altos y fuertes.
Dentición de desarrollo idéntica a la actual aunque con dientes más similares a H. erectus.
Cavidad craneal grande (+1.000 cm3). Cerebro más pequeño que el actual y con menos circunvoluciones. Forma de la mandíbula similar a formas modernas.
Canibalismo frecuente y presente en los huesos encontrados.
Vídeo donde podéis un poco del yacimiento sin ir a Burgos
Si os gusta la radio, Memorias de un tambor es uno de los mejores podcast de historia en español. Este programa se dedica a Atapuerca y el H. antecessor
4.5.6. Homo heidelbergensis (500.000 a. - 250.000 a.)
Su nombre científico deriva de la proximidad de la ciudad de Heidelberg (Alemania) al lugar donde fueron hallados los primeros fósiles, lo que subraya el hecho de que se trata de los primeros Homo que alcanzaron las estepas del centro y norte de Eurasia.
Su anatomía ha sido descrita en gran parte por los restos encontrados en el yacimiento de la Sima de los Huesos en Atapuerca. Eran individuos altos que tenían 1,75 m de estatura y muy fuertes (llegarían a 105 kg), de grandes cráneos que median 1350 cm³, muy aplanados con relación a los del hombre actual, con mandíbulas salientes y gran abertura nasal. Se trata de la primera especie humana en la que es posible detectar indicios de una mentalidad simbólica (se han encontrado restos de pinturas).
En Terra Amata (Francia), se encontraron restos de refugios construidos durante la época de H. heidelbergensis. Estos refugios tenían una forma ovalada, con veinticinco pies de largo y veinte de ancho. Dentro del refugio se encontraron restos de ceniza.El uso del fuego está acreditado con múltiples restos. Esto permite conservar y esterilizar los alimentos, conseguir calor y luz y permitir una vida nocturna activa (lo que incrementa la socialización en todos los sentidos). También permite mejoras tecnológicas en herramientas, armas, pinturas etc etc.
Muestra signos evidentes de comunicación hablada. Tanto por una elevada cefalización en las áreas de Broca y Wernicke como por la estructura de su laringe.
4.5.7. Homo rhodesiensis (600.000 - 160.000 a.)
Se considera que su distribución es exclusivamente africana.
Su capacidad craneal es elevada (1280-1325 cm3). Tiene rasgos comunes a muchas especies como H. erectus, H. ergaster y H antecessor; también otros que lo relacionan con H. heidelbergensis y otros que apuntan al H. sapiens. En general podemos citar todo el desarrollo de H. heidelbergensis en cuanto a herramientas, fuego y conducta social.
Hay autores que opinan que H. antecessor, H.heidelbergensis y H. rhodesiensis son una única especie (H heidelbergensis en sentido amplio) de la que derivarían H. neanderthalensis en Europa y H. sapiens en África. La otra teoría, que hemos seguido nosotros, es que hay una rama preneandertal en Europa y otra rama presapiens a partir de H. antecessor.
4.5.8. Homo neanderthalensis
Los neandertales son una especie extinta del género Homo que habitó en Europa, Próximo Oriente, Oriente Medio y Asia Central durante el final del Pleistoceno medio y casi todo el superior. Esta especie evoluciona en Europa a partir del H. heidelbergensis. Su evolución se debe a la glaciación de Mindel (400-350.000 a.) que obliga a H. heidelbergensis a emigrar a la península ibérica, italiana y griega huyendo de la extensión de los glaciares por toda Europa central. Esto provoca que estas poblaciones se reduzcan y aíslen mucho lo que favorece la especiación” (como ya hemos estudiado).
Algunos autores lo consideran como una subespecie de Homo sapiens, y se suelen referir a dicha subespecie como Homo sapiens neanderthalensis (debido a que los neandertales y los sapiens han hibridado de forma fértil en al menos dos lugares (Próximo Oriente y Europa occidental).
Presentaban numerosas adaptaciones derivadas de una evolución en climas fríos: baja estatura, complexión robusta, nariz amplia (rasgos que podemos observar en poblaciones árticas de H. sapiens). También presentaban despigmentación de la piel (piel clara) y el pelo (los ejemplares encontrados eran pelirrojos).
Anatómicamente, los neandertales eran más robustos que el humano moderno, con un tórax y cadera anchos y extremidades cortas. Su estatura estaba alrededor de 1,65-,170 m (un poco más bajo que H. sapiens) y un esqueleto y musculatura eran muy potentes (alcanzaban fácilmente los 100 Kg y sus huesos eran muy difíciles de romper). Probablemente no eran buenos corredores de larga distancia pero si grandes marchadores y rápidos en carreras cortas de persecución/huida.
El cráneo se caracteriza por su doble arco superciliar, frente huidiza, la ausencia de mentón y una capacidad craneal media más grande, 1550 cm3, que la de Homo sapiens.
Los neandertales eran omnívoros y explotaban una amplia variedad de alimentos pesqueros, mariscos, vegetales, etc. También eran grandes cazadores y depredaban mamuts y todo tipo de mega y mesofauna. Se han encontrado heridas en los fósiles de los animales que demuestran el uso de lanzas a distancia y una caza muy organizada y coordinada.
Presentan canibalismo de tipo ritual (probablemente asociado a rituales religiosos de enterramiento) y, en algunos casos, alimenticio.
Se ha discutido mucho sobre su dominio de un lenguaje articulado, complejo y simbólico como el de sapiens. Los recientes descubrimientos de que tenían un hueso hiodes igual que el de sapiens, sus técnicas de caza, sociales, herramientas, su desarrollo cerebral y sus genes parecen indicar claramente que tenían un lenguaje articulado y simbólico. Las simulaciones hechas por ordenador también lo muestran claramente.
El tipo de herramientas líticas que se han encontrado, y a las que se les asocia, se adscriben a la denominada cultura Musteriense, característica del Paleolítico medio. En los últimos años de existencia de los neandertales, aparecen en el registro arqueológico herramientas diferentes que se incluyen en la cultura Châtelperroniense, que algunos autores atribuyen al Homo sapiens (aunque puede ser por los prejuicios que se han ido desterrando sobre los neandertales).
Herramientas de piedra (Musteriense)
Herramientas de piedra (Châtelperroniense)
Estas herramientas de piedra se tallaban usando martillos de percusión blandos (hueso o madera). Los neandertantes nunca fueron capaces de producir una tecnología tan elaborada como desarrollaron los sapiens y puede que se debiera a que las falanges de sus manos eran más cortas (lo que dificultaría enormemente elaborar objetos pequeños con precisión).
Antiguamente se les consideraba incapaces de un pensamiento simbólico pero los últimos años se han descubierto pinturas rupestres en España de hace 65.000 a. (20.000 a. antes de la llegada deH. sapiens), adornos, máscaras, …”
Garras de águila blanca presumiblemente para un colgante o collar
Diversos ornamentos corporales de hueso y conchas
Se han encontrado plumas, conchas decoradas y que contenían pigmentos (lo que hace pensar en que se pintaban o maquillaban) y muy anteriores a su contacto con sapiens (antes se pensaba que estaban “copiando” a los sapiens).
También se ha descubierto que sabían cocinar (por los restos de comida encontrados), que cuidaban a los ancianos y enfermos (se han encontrado fósiles de individuos que arrastraban lesiones y enfermedades que no les hubieran permitido sobrevivir sin cuidados) e incluso el uso de medicinas (como hongos que producen antibióticos en infecciones).
Usaban las cuevas mucho antes que los sapiens y tenían lugares específicos para cada tipo de uso: dormitorio, despensa, etc. En Gibraltar se han encontrado algunos de los establecimientos más importantes y mejor estudiados de Europa. El fuego, tan importante para ellos, siempre tenía una posición central en sus asentamientos.
Por el desgaste diferente en dientes y huesos parece que tenían un reparto del trabajo por sexos (como ocurre en las sociedades recolectoras-cazadoras sapiens.
También ha quedado claro que pensaban que existía la vida después de la muerte y tenían algún tipo de creencias religiosas. Enterraban de forma ritual a sus muertos y frecuentemente les dejaban ofrendas, flores y herramientas que pudieran servirles en la otra vida. También que las antiguas teorías de que cuidaban menos a sus hijos sean probablemente falsas dado que los enterramientos de individuos jóvenes suelen ser más cuidados y elaborados.
Enterramiento en una cueva artificial. Se les colocaba en posición fetal con diversos utensilios y ofrendas
Se desconocen las causas exactas de su extinción. Probablemente fue la conjunción del desplazamiento por parte del H. sapiens a las zonas más hostiles unido al enfriamiento del clima y que las pequeñas poblaciones fueron quedando cada vez más aisladas y, por tanto, con mayores niveles de endogamia, lo que acabó con la extinción de la especie. Tampoco hay que descartar el hecho de que pueden haber sido absorbidos por H. sapiens que llegaba en números mucho mayores en oleadas desde África; al reproducirse ambos el ADN de los neandertales fue quedando cada vez más diluido (hoy en día los H. sapiens europeos tenemos entre un 1,5-2% de ADN neandertal).
4.5.9. Homo sapiens (130.000a.-...)
Aparece como tal en África oriental hace alrededor de 165.000 años. Posteriormente se diseminó por el continente africano y salió en dos grandes oleadas: una más antigua que colonizó Asia y Oceanía (donde hibrido parcialmente con una especie que no hemos estudiado al haber muy pocos restos aparte de los genéticos: los devisonianos) y una mucho más reciente que colonizo Europa (donde hibridó parcialmente con los neandertales).
Tiene una anatomía igual a la nuestra y una gran capacidad cerebral (1350 cm3), con una frente erguida, una cara plana y más reducida, presencia de mentón en la mandíbula, una dentición algo menor y un esqueleto poscraneal algo más grácil.
Mantiene un notorio dimorfismo sexual en el nivel anatómico, por ejemplo, la talla media actual entre los varones caucásicos (si crecen bien nutridos y con poco estrés) hacia los 21 años es de 1,75 m, la talla media de las mujeres caucásicas en iguales condiciones es de 1,62 m, y los pesos promedios respectivos son de 75 kg y 61 kg.
No vamos a estudiar detalladamente su anatomía (ya lo hicisteis en 3º) ni su historia o cultura (ya lo hicisteis en historia) pero sí cabe destacar que son un poco más altos y ligeros que los neandertales con lo que sus desplazamientos consumirían menos energía.
Su nivel social y tecnológico inicial no es especial pero sí van a ir desarrollando conductas y tecnologías más complejas y ricas que las especies anteriores (que acabaron desembocando en la gran revolución neolítica (que habéis estudiado en historia).
4.6. Proceso de hominización
Llamamos proceso de hominización al proceso evolutivo que condujo a que el hombre adquiriera las características que lo definen. La mayoría de estos caracteres tenían una retroalimentación positiva y coevolucionaban en la mismo sentido. Por ejemplo cuánto más inteligente era la especie más necesitaba el lenguaje para comunicarse y cuánto mejor y más complejo era su lenguaje más necesitaba ser más inteligente. Algo equivalente ocurre con el desarrollo de la habilidad manual y la inteligencia.
Las principales características que han hecho a nuestra especie lo que es son:
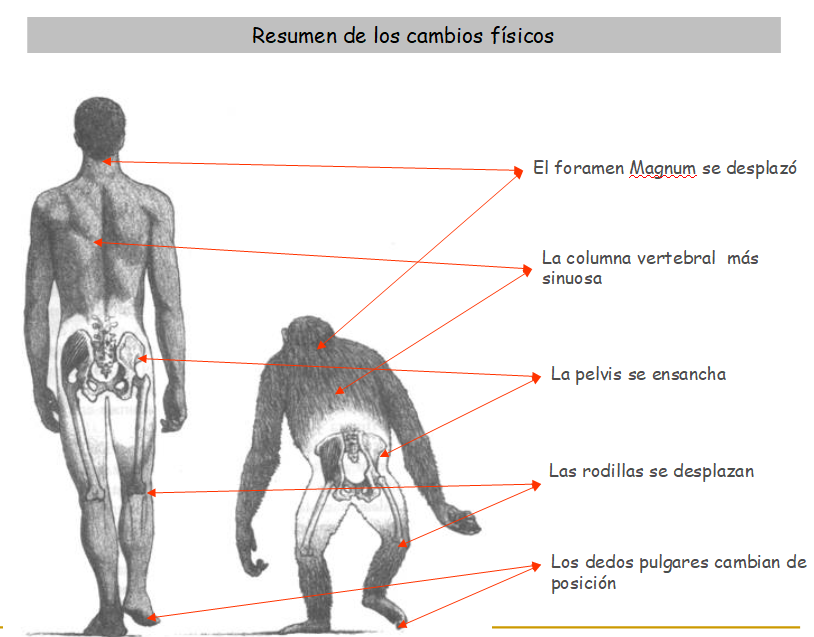
4.6.1. La bipedestación
La bipedestación es muy útil en la sabana (medio donde evolucionó el hombre) pues permite ver lejos y además supone un menor gasto energético comparado con la locomoción cuadrúpeda.
Esta forma de andar supone:
a) estrechamiento de la pelvis (lo que ocasiona graves problemas de cara al parto).
b) curvatura en la columna vertebral en forma de doble S para equilibrar el peso.
c) un pulgar no oponible y unos dedos no curvados en los pies para originar la planta del pie.
d) un alargamiento de las piernas y un aumento en el ángulo de las rodillas.
e) un desplazamiento del foramen magnum que conlleva el progresivo acortamiento de la cara.
La bipedestación trae un beneficio añadido que se revelará de gran importancia que es la liberación de las manos durante los desplazamientos. Esto permitirá que se desarrollen como apéndices exclusivamente manipulativos y su destreza pueda aumentar.
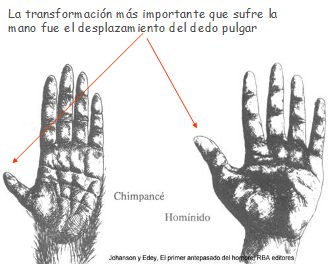
4.6.2. La liberación de las manos
La existencia de un pulgar oponible y la posibilidad de extensión y flexión de la mano posibilitaba coger y usar objetos. Cuando dejan de usarse para moverse pueden evolucionar hasta formar una pinza de enorme precisión. Su coevolución con el sistema nervioso y la inteligencia permite la fabricación de objetos con múltiples usos.
La palma de la mano se va a acortar volviéndose prácticamente cuadrada y los dedos se van a alargar volviéndose más precisos y móviles pudiendo llegar a tocar con la yema del pulgar la yema de todos los demás dedos.
El uso de herramientas fabricadas y usadas con precisión va a permitir a los humanos acceder a alimentos antes inaccesibles (tuétano de los huesos, resto carne adheridos huesos) y va a traer un gran beneficio añadido que es la reducción de su dentadura. Al cortar y machacar los alimentos con herramientas sus piezas dentarias y sus músculos masticadores no necesitan tanta fuerza y van a ir disminuyendo su tamaño.
6.6.3. La cefalización
Como hemos visto en el punto anterior hay un aumento progresivo de la capacidad craneana durante toda la evolución del género Homo.
También hay un aumento en la complejidad de la corteza cerebral aumentando las circunvoluciones cerebrales (como se ha visto estudiando los moldes del interior de los cráneos fósiles).
El aumento del tamaño del cráneo unido al estrechamiento de la pelvis genera partos dolorosos y difíciles. Debido a esto el cerebro del bebé todavía se desarrolla durante unos meses después del nacimiento.
6.6.4. El desarrollo del lenguaje
Con el desarrollo de la inteligencia y la vida en comunidades el desarrollo de un lenguaje capaz de establecer una comunicación compleja y eficaz se convierte en una gran ventaja evolutiva.
Las áreas de Broca y Wernicke de la corteza cerebral van a desarrollarse progresivamente ya desde los primeros miembros del género Homo.
De forma paralela (aunque más lenta) la disminución de la mandíbula (que ya hemos estudiado), las piezas dentales y los músculos masticatorios va a permitir un descenso de la laringe y que músculos especializados se dediquen a elaborar sonidos articulados y complejos. La aparición de las cuerdas vocales y el hueso hioides facilitan un lenguaje como el que tenemos.
6.6.5. Neotenia
La neotenia o inmadurez biológica se produce en la evolución humana para permitir que el gran cráneo de los bebés pueda pasar por el canal del parto. Como consecuencia de este adelanto del parto los bebés humanos nacen muy poco formados e indefensos.
También se ha potenciado evolutivamente un retraso madurativo que hace que su llegada a la edad reproductiva sea muy lenta. Esto permite un mayor tiempo de aprendizaje así como el establecimiento de relaciones sociales y afectivas mucho más profundas entre los miembros del grupo.



























































































































































No hay comentarios:
Publicar un comentario