Los lípidos constituyen un grupo de sustancias químicamente muy heterogéneo que no se caracteriza, como otras biomoléculas, por la posesión de un determinado conjunto de grupos funcionales. Aunque químicamente heterogéneos, todos presenten un denominador común estructural: la totalidad, o al menos una parte significativa, de su molécula es de naturaleza hidrocarbonada, y por lo tanto apolar.

Por ello, resulta mucho más conveniente identificarlos sobre la base de sus propiedades físicas: se caracterizan por ser poco o nada solubles en agua y, por el contrario, muy solubles en disolventes orgánicos no polares (éter, cloroformo, benceno) con los que pueden establecer interacciones hidrofóbicas (fuerzas de Van der Waals). Son poco densos y suelen flotar sobre el agua.
Otras de sus características físicas es que son untuosos al tacto y suelen tener un brillo característico (graso).
Según su composición química, los lípidos se clasifican en lípidos simples (C, H, O) y lípidos complejos (C, H, O, N, P). Sin embargo, la clasificación que solemos usar los divide en:
a) Lípidos saponificables; aquellos que contienen ácidos grasos unidos a un alcohol (y que por tanto pueden hidrolizarse).
b) Lípidos insaponificables; aquellos que no contienen ácidos grasos y, por lo tanto, no pueden hidrolizarse. Su monómero teórico es el isopreno.

1. Ácidos grasos
Los ácidos grasos son compuestos orgánicos que poseen un grupo funcional carboxilo y una cadena hidrocarbonada larga con un número par de carbonos. La fórmula general es por lo tanto: CH3 – (CH2)n - COOH, siendo n un número par entre 10 y 22 (los más comunes tienen entre 16 y 18 carbonos). El número de carbonos es siempre par porque se sintetizan a partir de una molécula con dos carbonos (como veremos más adelante).

Los ácidos grasos son moléculas anfipáticas: tienen una parte polar (grupo carboxilo) y una gran cadena apolar (cadena hidrocarbonada). Esto va a provocar unos comportamientos especiales en medio acuoso que veremos más adelante.

Los ácidos grasos no suelen encontrarse libres en la naturaleza como tales, sino que forman parte de las macromoléculas de los lípidos. Existen dos tipos principales de ácidos grasos:
a) Ácidos grasos saturados; no poseen dobles enlaces,
b) Ácidos grasos insaturados; poseen uno o más dobles enlaces a lo largo de su cadena hidrocarbonada. La existencia de dobles enlaces implica la existencia de isómeros geométricos (cis-trans) según sea la disposición de los sustituyentes a ambos lados del doble enlace. La mayoría de los ácidos grasos insaturados que existen en la naturaleza presentan configuración cis.
La geometría tetraédrica de los orbitales del carbono determina que las cadenas hidrocarbonadas de los ácidos grasos adopten una característica disposición en zig-zag. Sin embargo, los ácidos grasos saturados e insaturados difieren significativamente en cuanto a la disposición espacial de sus cadenas hidrocarbonadas.
En los saturados, aunque la libre rotación de los sustituyentes alrededor de los enlaces sencillos proporciona una gran flexibilidad a la cadena, la conformación más estable es aquella en la que dicha cadena se encuentra lo más extendida posible, minimizando así las interacciones repulsivas entre átomos vecinos.

En los insaturados, por otra parte, la tendencia de la cadena hidrocarbonada a adoptar la conformación más extendida se ve limitada por la rigidez de los dobles enlaces, que impide que los distintos sustituyentes de los carbonos implicados en ellos puedan rotar a su alrededor. Esto determina la aparición de cambios de orientación en la dirección de la cadena hidrocarbonada de los ácidos grasos insaturados, no pudiendo adoptar ésta una conformación totalmente extendida. En concreto, en los ácidos grasos cis-monoinsaturados, que son los más abundantes, la cadena presenta dos tramos rectilíneos, separados por un doble enlace, que forman entre sí un ángulo de unos 120º.


Los ácidos grasos cis-poliinsaturados presentan estructuras complejas con varios tramos rectilíneos separados por dobles enlaces.

Los dobles enlaces trans (muy raros en la naturaleza) apenas determinan una ligera sinuosidad en la cadena sin que ello suponga un cambio significativo en la orientación de la misma: los ácidos grasos trans-insaturados presentan conformaciones espaciales muy similares a las de los saturados. Estas diferencias en cuanto a la conformación espacial de los distintos tipos de ácidos grasos influyen considerablemente en sus propiedades físicas y tienen notables implicaciones biológicas.


Algunos ácidos grasos poliinsaturados son esenciales, es decir, nuestro cuerpo no puede sintetizarlos, dado que la maquinaria celular es incapaz de colocar dobles enlaces en dichas posiciones, por lo que debemos ingerirlos en la dieta. Los ácidos grasos esenciales más conocidos son el linoleico (omega-6) y α-linolénico (omega-3), de origen vegetal; y el araquidónico (omega-6), de origen animal, que se encuentra en la carne y huevos. Estos ácidos grasos son importantes para la síntesis de prostaglandinas, que veremos más adelante. La denominación en omega empieza por el final, pues omega (Ω) es la última letra del alfabeto griego. Por ej., un ácido omega-6 es el que presenta el primer doble enlace en el carbono 6 empezando a contar por el final de la cadena hidrocarbonada.

1.1 Propiedades físicas de los ácidos grasos
Las propiedades físicas de los ácidos grasos vienen determinadas en gran medida por la longitud y grado de insaturación de su cadena hidrocarbonada. Entre estas propiedades cabe destacar, por su importancia biológica, dos de ellas:
a) Punto de fusión; el punto de fusión de los ácidos grasos aumenta gradualmente con la longitud de su cadena hidrocarbonada. Cuando los ácidos grasos se solidifican sus moléculas se empaquetan formando un retículo regular en el que cada una de ellas se encuentra unida a sus vecinas mediante interacciones de Van der Waals entre las respectivas cadenas hidrocarbonadas. Cuanto más largas sean dichas cadenas mayor será el número de interacciones que se podrán establecer entre ellas y, por lo tanto, más cantidad de energía térmica habrá que emplear paran romperlas y pasar así del estado sólido al estado líquido, es decir, mayor será el punto de fusión.
Los ácidos grasos saturados tienen puntos de fusión significativamente mayores que los insaturados de igual número de átomos de carbono. Esto se debe a que la conformación extendida de los ácidos grasos saturados permite que sus moléculas se empaqueten muy estrechamente estableciéndose interacciones de Van der Waals todo a lo largo de sus cadenas hidrocarbonadas; por el contrario, los cambios de orientación existentes en las cadenas hidrocarbonadas de los ácidos grasos cis-insaturados impiden que sus moléculas se empaqueten tan estrechamente dificultando la formación de interacciones de Van der Waals.


Así, al existir entre las cadenas hidrocarbonadas de los ácidos grasos saturados un mayor número de interacciones de Van der Waals, la energía térmica necesaria para romper estas interacciones es mayor, lo que se traduce en un mayor punto de fusión de los ácidos grasos saturados (y trans-insaturados).
El punto de fusión de los ácidos grasos determina el de los lípidos que los contienen. Es muy importante que determinadas estructuras lipídicas, como las membranas celulares, permanezcan fluidas, y por ello los distintos tipos de organismos deben regular la composición en ácidos grasos de sus lípidos constituyentes. Así, en los vegetales y los animales poiquilotermos, que no mantienen una temperatura corporal constante, abundan los ácidos grasos insaturados, que tienen un punto de fusión bajo, mientras que los animales homeotermos, que mantienen una temperatura corporal constante y elevada, pueden recurrir en mayor medida a los ácidos grasos saturados sin correr el riesgo de que sus membranas "cristalicen" cuando la temperatura exterior es muy baja.
b) Comportamiento en disolución; los ácidos grasos son sustancias anfipáticas: el grupo carboxilo, que a pH=7 se encuentra ionizado, es netamente polar, mientras que la cadena hidrocarbonada es totalmente no polar. Por lo tanto, en medio acuoso los ácidos grasos tenderán a formar monocapas, bicapas, micelas y otras estructuras afines.

2.1 Grasas o triacilglicéridos
Los acilglicéridos son ésteres de la glicerina, un polialcohol de tres átomos de carbono, con los ácidos grasos. La glicerina (o glicerol) puede considerarse como un azúcar-alcohol que deriva biológicamente de la dihidroxiacetona (una cetotriosa); sus tres grupos hidroxilo pueden reaccionar con uno, con dos o con tres ácidos grasos para dar lugar respectivamente a los monoacilglicéridos, diacilglicéridos y triacilglicéridos.

Cuando la glicerina reacciona con tres ácidos grasos para dar lugar a un triacilglicérido se liberan tres moléculas de agua.

Esta reacción de esterificación es reversible en determinadas condiciones, es decir, los triacilglicéridos pueden sufrir hidrólisis cuando reaccionan con el agua para rendir de nuevo la glicerina y los ácidos grasos libres, tal y como sucede durante la digestión de las grasas en el intestino delgado por acción de la lipasa pancreática.
Por otra parte, los triacilglicéridos pueden sufrir una saponificación cuando reaccionan con álcalis (bases) como el hidróxido sódico (NaOH) para dar lugar a la glicerina libre y a las correspondientes sales sódicas de los ácidos grasos que se conocen con el nombre de jabones. Esta reacción no es exclusiva de los triacilglicéridos, sino que la sufren en general todos los lípidos que contienen ácidos grasos unidos, mediante enlace éster o similar, a otro componente; por ello, el carácter saponificable o no de los distintos tipos de lípidos se utiliza como criterio para clasificarlos. La reacción de saponificación se utiliza industrialmente en la fabricación de jabones.



Las moléculas de jabón son sustancias anfipáticas cuya cabeza polar (representada con un círculo) se une a otras sustancias polares como el agua y cuya cola apolar (representada con una línea irregular) atrae sustancias apolares como las grasas. De esta manera, el jabón lava porque las sustancias apolares son rodeadas por la parte apolar del jabón, y el agua, al arrastrar el jabón (se une a la parte polar del jabón), se lleva la suciedad.

Los triacilglicéridos pueden ser simples, si contienen un sólo tipo de ácido graso, o mixtos, si contienen más de un tipo. Los triacilglicéridos naturales suelen ser mezclas complejas de triacilglicéridos simples y mixtos. Por otra parte, los triacilglicéridos ricos en ácidos grasos saturados se encuentran en estado sólido a temperatura ambiente y se denominan sebos o tocinos, mientras que los ricos en ácidos grasos insatura-dos permanecen líquidos a temperatura ambiente y se denominan aceites.
La polaridad típica de los grupos hidroxilo de la glicerina y carboxilo de los ácidos grasos desaparece por completo cuando éstos reaccionan para formar un enlace éster. Por ello, aunque la glicerina es una sustancia polar y los ácidos grasos son sustancias anfipáticas, los triacilglicéridos son totalmente apolares y por lo tanto insolubles en agua. Esta insolubilidad se pone claramente de manifiesto en las mezclas agua-aceite, que presentan dos fases completamente separadas.
2.1.1 Funciones de las grasas o triacilglicéridos
La principal función de los triacilglicéridos en las células vivas es de reserva energética, constituyen depósitos de combustible metabólico rico en energía al que la célula puede recurrir en períodos en los que escasean los nutrientes procedentes de su entorno. Se almacenan en forma de gotículas microscópicas que forman un fase separada del citosol acuoso circundante.
Como sustancias de reserva los triacilglicéridos presentan dos ventajas significativas con respecto a los polisacáridos como el almidón o el glucógeno, que comparten con ellos esta función. En primer lugar, por ser sustancias altamente reducidas (o hidrogenadas), su oxidación libera más del doble de energía que una cantidad equivalente de glúcidos (9 Kcal/g frente a 4 Kcal/g), que presentan un grado de oxidación mayor y por lo tanto no son tan ricos en energía. En segundo lugar, por ser sustancias hidrofóbicas, pueden almacenarse en forma anhidra, sin que el organismo tenga que soportar el peso adicional del agua de hidratación de los polisacáridos, mucho más hidrofílicos.
Por esta razón, los animales, que dependen en gran medida de la locomoción para desarrollar sus funciones vitales, recurren preferentemente a los lípidos como material de reserva, ya que la misma cantidad de energía en forma de polisacáridos hidratados dificultaría sus movimientos debido al exceso de peso; los vegetales, por ser estáticos, no tienen este problema, por lo que pueden recurrir en mayor medida a los polisacáridos para almacenar su combustible metabólico.
En algunos animales los triacilglicéridos no actúan sólo como sustancias de reserva energética, sino que, por encontrarse almacenados preferentemente en el panículo adiposo existente bajo la piel, desempeñan también la función de aislante térmico para proteger al organismo de las bajas temperaturas. También es reseñable su función de amortiguadores mecánicos en zonas como las articulaciones o alrededor de los órganos vitales.
Los ceras son ésteres de los ácidos grasos con alcoholes monohidroxílicos de cadena larga (16 a 30 átomos de carbono), que también se denominan alcoholes grasos. El éster formado por el ácido palmítico (16C) y el triacontanol (alcohol graso de 30C) es el componente principal de la cera que fabrican las abejas.


Al igual que los triacilglicéridos, los céridos son sustancias netamente hidrofóbicas (tienen dos cadenas hidrocarbonadas muy largas y apolares) y por lo tanto insolubles en agua. Esta insolubilidad en agua junto con su elevada consistencia constituyen la base físico-química de su principal función biológica que consiste en actuar como sustancias impermeabilizantes. Así, ciertas glándulas de la piel de los vertebrados secretan ceras para proteger el pelo y la piel manteniéndolos flexibles, lubricados e impermeables; los pájaros, especialmente las aves acuáticas, secretan ceras gracias a las cuales sus plumas pueden repeler el agua; en muchas plantas, sobre todo las que viven en ambientes secos, las hojas y los frutos están protegidas contra la excesiva evaporación de agua por películas céreas que les dan además un aspecto brillante característico.
Las ceras naturales tienen diversas aplicaciones en las industrias farmacéutica y cosmética.
2.3 Fosfolípidos o fosfoglicéridos
Los fosfoglicéridos, son un grupo de lípidos con un denominador estructural común que es la molécula de ácido fosfatídico. El ácido fosfatídico está formado por una molécula de glicerina, dos ácidos grasos y una molécula de ácido fosfórico.
Los grupos hidroxilo de los átomos de carbono 1 y 2 de la glicerina están unidos mediante enlace éster a los dos ácidos grasos (normalmente el esteárico, saturado, con el C1 y el oleico, insaturado, con el C2), el hidroxilo del carbono 3 está unido, también mediante enlace éster, al ácido fosfórico. El ácido fosfatídico apenas se encuentra como tal en la naturaleza, sino unido a través de su molécula de ácido fosfórico a diferentes compuestos de naturaleza polar, dando así lugar a los diferentes fosfoglicéridos. La unión entre el ácido fosfatídico y el compuesto polar también es de tipo éster.
Los fosfoglicéridos presentan una cierta similitud con los triacilglicéridos: se podría concebir un fosfoglicérido como un triacilglicérido en el que uno de los ácidos grasos ha sido sustituido por una molécula de ácido fosfórico y un compuesto polar unido a él. Los compuestos polares que forman parte de los fosfoglicéridos son de naturaleza variada; entre ellos cabe citar algunas bases nitrogenadas como la colina y la etanolamina, algún aminoácido como la serina, o el azúcar-alcohol cíclico inositol. Todos ellos tienen en común su carácter acusadamente polar.



Los fosfoglicéridos son sustancias anfipáticas: tienen en su molécula una parte no polar, representada por las cadenas hidrocarbonadas de los dos ácidos grasos y el esqueleto de la glicerina, y una parte polar formada por el ácido fosfórico y el compuesto polar. Como hemos visto en la ilustración superior, es habitual representar a los fosfoglicéridos mediante una "cabeza" polar y dos "colas" no polares.
El carácter anfipático de los fosfoglicéridos constituye la base físico-química de su principal función biológica, que es la de ser componentes esenciales de las membranas celulares. Otras sustancias anfipáticas más simples, como los ácidos grasos, son de forma aproximadamente cónica, por lo que tienden a formar micelas en medio acuoso; sin embargo, los fosfoglicéridos, por tener dos "colas" hidrocarbonadas, son de forma aproximadamente cilíndrica, y por ello tienden a formar en medio acuoso estructuras más complejas como las bicapas, las cuales a su vez pueden doblarse sobre sí mismas dando lugar a estructuras cerradas, con un compartimiento acuoso interior, que se denominan liposomas.

La estructura de las membranas celulares está basada en una bicapa formada por lípidos con carácter anfipático. Es probable que las primeras células que existieron en el océano primitivo se parecieran mucho a los liposomas, estando aisladas de su entorno por una simple bicapa lipídica que posteriormente evolucionó dando lugar a las actuales membranas.

2.4 Esfingolípidos
Los esfingolípidos son un grupo de lípidos que contienen formando parte de su molécula al aminoalcohol graso llamado esfingosina. El grupo amino de la esfingosina puede reaccionar con el grupo carboxilo de un ácido graso para formar entre ellos un enlace tipo amida dando lugar a un compuesto denominado ceramida, que es la base estructural común de todos los esfingolípidos de manera análoga a como el ácido fosfatídico lo es de los fosfoglicéridos.
Los distintos tipos de esfingolípidos se obtienen mediante la unión de la ceramida con distintos tipos de compuestos de naturaleza polar. Hay dos tipos principales de esfingolípidos:
a) Esfingofosfátidos; también llamados esfingomielinas. Se obtienen al unirse la ceramida mediante enlace éster a una molécula de ácido fosfórico y ésta a su vez, también mediante enlace éster, a una base nitrogenada de carácter polar como la colina o la etanolamina, también presentes en los fosfoglicéridos.


b) Glucoesfingolípidos; se obtienen al unirse la ceramida a un azúcar mediante enlace glucosídico. Así pues, la unión de la ceramida al compuesto polar se realiza directamente y no a través de una molécula de ácido fosfórico como en los esfingofosfátidos; los glucoesfingolípidos no contienen fósforo. Se distinguen dos tipos de glucoesfingolípidos según la naturaleza del azúcar componente: los cerebrósidos, que incorporan un azúcar sencillo tal como la glucosa o la galactosa, y los gangliósidos, que incorporan azúcares complejos formados por varios monosacáridos y derivados de éstos unidos entre sí.
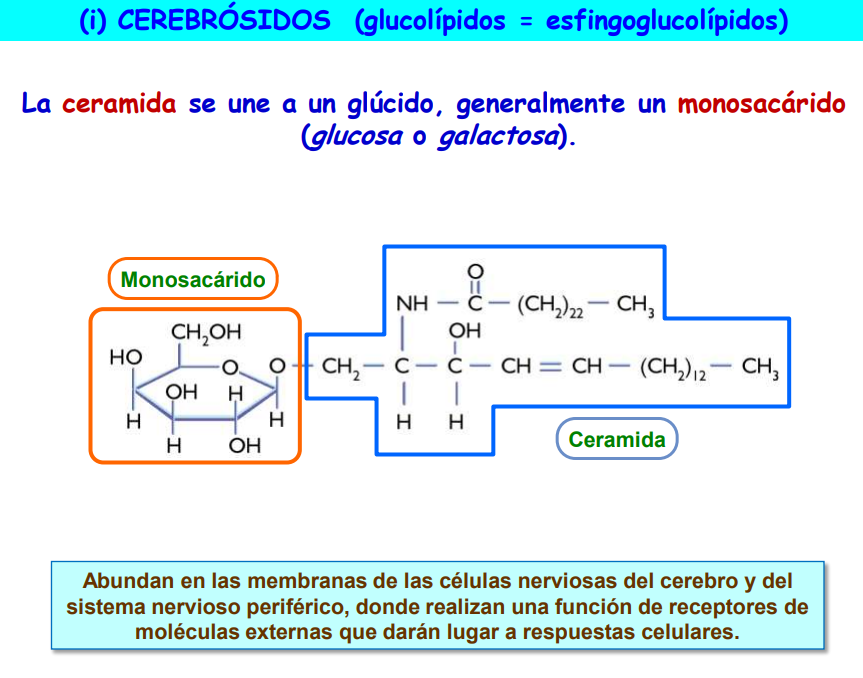

Los esfingolípidos presentan claras similitudes estructurales con los fosfoglicéridos: el papel del esqueleto de glicerina que éstos poseen es sustituido aquí por los tres primeros carbonos de la esfingosina; también presentan una "cabeza" polar (fosfato-base nitrogenada o azúcar) y dos "colas" no polares, que aquí corresponden a la cadena hidrocarbonada de la esfingosina y a la del ácido graso unido a ella.
Estas similitudes afectan también a sus propiedades: los esfingolípidos también presentan carácter anfipático y, por las mismas razones que los fosfoglicéridos, también son componentes esenciales de las membranas celulares. Abundan especialmente en las membranas de las células del tejido nervioso. Sin embargo, en los últimos años se ha descubierto que los esfingolípidos no son meros componentes estructurales de las membranas sino que tienen un importante papel en diversos procesos de reconocimiento de la superficie celular, por ejemplo, los determinantes de los grupos sanguíneos humanos (ABO) son glucoesfingolípidos. Por otra parte, diversas enfermedades graves que afectan al sistema nervioso están relacionadas con anomalías en el metabolismo de los esfingolípidos.
3. LÍPIDOS INSAPONIFICABLES
3.1 Terpenos
Los terpenos o isoprenoides son un grupo de lípidos que no contienen ácidos grasos y son por lo tanto no saponificables. Químicamente están formados por la polimerización de un hidrocarburo de 5 átomos de carbono llamado isopreno (2-metil-1,3-butadieno) y se clasifican según las unidades de isopreno que contienen (regla del isopreno) pero este nunca aparece como un producto natural y el verdadero precursor de los terpenos en el metabolismo es el ácido mevalónico (derivado del acetil Co-A).

Los terpenos existentes en la naturaleza contienen un número variable de unidades de isopreno polimerizadas. Generalmente la "cabeza" de cada unidad de isopreno está unida con la "cola" de la siguiente. Debido a su naturaleza hidrocarbonada son sustancias netamente hidrofóbicas y por lo tanto insolubles en agua.



Son especialmente abundantes en el mundo vegetal. Gracias a que presentan un sistema de dobles enlaces conjugados tienen la propiedad de absorber luz de diferentes longitudes de onda, por lo que pueden actuar:
a) Como pigmentos; algunos terpenos como los carotenos y xantofilas son los responsables de la coloración característica de muchos frutos y participan en la fotosíntesis absorbiendo determinadas longitudes de onda de la luz.
b) El fitol, un alcohol terpenoide, forma parte de la clorofila, el pigmento vegetal por excelencia.
c) Muchos aromas de origen vegetal, como el alcanfor, el mentol y el geraniol son de naturaleza terpenoide.
d) Productos vegetales de interés económico como el caucho.
e) La vitamina A, que deriva biológicamente de los carotenos, también es un terpeno y tiene un papel decisivo en la visión.
Otra posibilidad, al tener dobles enlaces conjugados, es participar en las cadenas de transporte de electrones y de energía, en este apartado se incluyen las vitaminas E y K, la E es un potente antioxidante y la K interviene en la coagulación sanguínea, así como plastocianina y plastiquinona que actúan en las membranas de los tilacoides en la fase luminosa de la fotosíntesis.
3.2 Esteroides
Los esteroides son un grupo de lípidos de estructura compleja que no contienen ácidos grasos y por lo tanto son no saponificables. Están relacionados estructuralmente con el hidrocarburo tetracíclico denominado ciclopentanoperhidrofenantreno. Biológicamente están relacionados con los terpenos, de los cuales derivan. Los diferentes esteroides se diferencian en la naturaleza y posición de diversos grupos funcionales, dobles enlaces y cadenas alifáticas lineales o ramificadas añadidas al anillo del ciclopentanoperhidrofenantreno.

Entre los esteroides se encuentran los esteroles, que poseen un grupo hidroxilo en el carbono 3 y un doble enlace C5=C6; son los más abundantes e incluyen muchas moléculas de interés biológico entre las que destaca el colesterol.
El colesterol desempeña importantes funciones biológicas en las células animales. En primer lugar, gracias a su moderado carácter anfipático, es un componente esencial de las membranas celulares, a las que proporciona fluidez intercalándose entre los demás lípidos de membrana para impedir que se agreguen demasiado y "solidifiquen" (sobre todo a bajas temperaturas). Por otra parte al situarse entre los fosfolípidos reduce su movilidad e impide que se separen con que disminuye la fluidez de la membrana. Por lo tanto es una molécula esencial para mantener la estabilidad de las membranas.
Se transporte por la sangre unido a lipoproteínas y puede depositarse en los vasos generando placas de ateroma. Durante muchos años se ha recomendado limitar su ingesta pero es sintetizado en el hígado y sus niveles dependen más de factores genéticos que de la dieta.
El colesterol es el principal esterol de los tejidos animales. En otros organismos existen esteroles semejantes, como el estigmasterol en las plantas y el ergosterol en los hongos. Con pocas excepciones las bacterias carecen de esteroles.

Los esteroles actúa como precursor de un amplio grupo de sustancias con actividades biológicas importantes entre las que cabe citar:
a) Hormonas sexuales como la testosterona (hormona sexual masculina), el estradiol (hormona sexual femenina), y la progesterona (hormona progestágena).
b) Hormonas adrenocorticales como la aldosterona y el cortisol, que regulan diferentes aspectos del metabolismo.
c) Ácidos biliares, principales componentes de la bilis, cuya función es emulsionar las grasas en el intestino delgado para facilitar la acción de las lipasas.
d) Vitamina D, que regula el metabolismo del fósforo y del calcio.


Como ya hemos visto, hay dos fuentes de colesterol en el cuerpo humano:
a) Colesterol exógeno: en alimentos ingeridos.
b) Colesterol endógeno: formado por el hígado.
Las lipoproteínas son partículas con un centro apolar, que incluye triacilgliceridos y ésteres de colesterol, y un revestimiento anfipático formado por fosfolípidos, colesterol no esterificado y las apolipoproteínas, que son lípidos asociados de forma no covalente con proteínas.


En los análisis de sangre para conocer el riesgo de sufrir un ataque coronario se realiza un perfil de lípidos que incluye:
a) Colesterol total.
b) Triglicéridos.
c) Lipoproteína de alta densidad (“colesterol” HDL); conocidas como colesterol bueno, recogen el colesterol no utilizado y lo devuelve al hígado para su almacenamiento o excreción al exterior a través de la bilis. Un nivel bajo de colesterol HDL (c-HDL) aumenta el riesgo de enfermedad cardiovascular.
d) Lipoproteína de baja densidad (“colesterol” LDL); también se conoce como “colesterol malo”. Las LDL llevan colesterol a los tejidos que lo necesitan para sus membranas (incluidas arterias) o para la síntesis de hormonas esteroideas. La mayor parte del colesterol en sangre es colesterol LDL (c-LDL). Cuanto mayor sea el nivel de colesterol LDL en sangre, mayor es el riesgo de enfermedad cardiovascular.
e) Lipoproteína de muy baja densidad (“colesterol” VLDL); contiene la mayor cantidad de triglicéridos. El VLDL se considera un tipo de “colesterol malo” puesto que ayuda a que el colesterol se acumule en las paredes de las arterias. Son las partículas que llevan los excesos de carbohidratos transformados en triglicéridos hacia el hígado.
La aterosclerosis es la formación de placas de ateroma en el interior de las arterias. Esta placa de ateroma contiene colesterol y otras grasas y supone un aumento del gasto cardiaco, que a su vez puede dar lugar a hipoxia y trombos.

Los trombos son pequeños coágulos de sangre que taponan los vasos y pueden dejar sin riego regiones del cuerpo. Si el trombo llega al corazón se produce una situación en la que el miocardio se ve privado de oxígeno, lo que provoca a menudo un gran dolor pectoral denominado angina de pecho.
Se produce un infarto de miocardio si una parte del miocardio se queda totalmente sin riego, generalmente por un trombo en la arteria coronaria. Es la causa principal de muerte en países desarrollados (70.000 personas al año en España).
3.3 Prostaglandinas
Son un grupo de lípidos que derivan de la ciclación de un ácido graso poliinsaturado de 20 átomos de carbono, el ácido araquidónico.

a) Algunas prostaglandinas estimulan la contracción del músculo liso del útero durante el parto o la menstruación.
b) Otras afectan al flujo sanguíneo
c) Al ciclo sueño-vigilia
d) Otras son las responsables de la fiebre y el dolor asociados a los procesos inflamatorios.
El conocido fármaco ácido acetilsalicílico (aspirina) actúa inhibiendo la síntesis de prostaglandinas, de ahí su acción analgésica y antipirética.
4. FUNCIONES GENERALES DE LOS LÍPIDOS
Los lípidos en los seres vivos desempeñan tres tipos de funciones:
a) Función energética; Aunque debido a su insolubilidad en agua, con la consiguiente dificultad para ser transportados en medio acuoso, los lípidos no pueden ser utilizados como combustible metabólico para un uso inmediato, constituyen (sobre todo los triacilglicéridos) un excelente almacén de combustible metabólico a largo plazo.
Esto es debido a que se conservan deshidratados (mucho menos peso) y aportan más energía que el resto de biomoléculas (9 kcal/g frente a 4 kcal/g).
b) Funciones estructurales; algunos tipos de lípidos (fosfoglicéridos, esfingolípidos y colesterol) son componentes esenciales de las membranas celulares. Otros como las ceras desempeñan funciones de protección y revestimiento de determinadas superficies, o de aislamiento térmico del organismo, como los triacilglicéridos almacenados en el tejido adiposo.
c) Funciones dinámicas; los lípidos más abundantes desempeñan en las células papeles relativamente "pasivos" como servir de combustible o formar parte de las membranas. Sin embargo, otros lípidos más escasos realizan importantes funciones de control y regulación del metabolismo celular. Así, algunas vitaminas y coenzimas son de naturaleza lipídica, como lo son también algunas hormonas, pigmentos fotosintéticos y otras biomoléculas de especial relevancia para la vida de las células.
No hay comentarios:
Publicar un comentario