1. El núcleo celular
El núcleo es la estructura más destacada de la célula eucarionte, tanto por su morfología como por sus funciones. colocación lateral en el caso de las células vegetales, o bien esférico y de colocación central en el caso de las células animales.
Suele ser único, y su tamaño es muy variable (por término medio el 10% del
volumen celular), siendo mayor en células que son muy activas, como por ejemplo las de los tejidos secretores o reproductores.
Existe una relación núcleo-plasmática que viene expresada por la siguiente expresión:
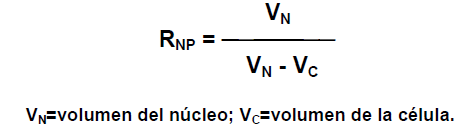
Esta relación es constante en un mismo tipo celular y cuando se altera, es cuando se induce el proceso de división celular.
El núcleo tiene tres funciones primarias (todas ellas relacionadas con su contenido de ADN):
a) Almacenar la información genética en el ADN.
b) Recuperar la información almacenada en el ADN en la forma de ARN.
c) Ejecutar, dirigir y regular las actividades citoplasmáticas, a través del producto de la expresión de los genes: las proteínas.
En el núcleo se localizan los procesos a través de lo cuales se llevan a cabo dichas funciones. Estos procesos son:
a) La duplicación del ADN y su ensamblado con proteínas (histonas) para formar la cromatina.
b) La transcripción de los genes a ARN y el procesamiento de éstos a sus formas maduras, muchas de las cuales son transportadas al citoplasma para su traducción.
c) La regulación de la expresión genética.
1.1 Estructura del núcleo interfásico
El núcleo está rodeado por la envoltura nuclear, una doble membrana interrumpida por numerosos poros nucleares. Los poros actúan como una compuerta selectiva a través de la cual ciertas proteínas ingresan desde el citoplasma, como también permiten la salida de los distintos ARN y sus proteínas asociadas.
La envoltura nuclear es sostenida desde el exterior por una red de filamentos intermedios dependientes del citoesqueleto, mientras que la lámina nuclear, la cual se localiza adyacente a la superficie interna de la envoltura nuclear, provee soporte interno.
El núcleo también tiene un nucleoplasma, en el cual están disueltos sus solutos y un esqueleto filamentoso, la matriz nuclear la cual provee soporte a los cromosomas y a los grandes complejos proteicos que intervienen en la replicación y transcripción del ADN.
Los cromosomas aparecen ocupando lugares específicos. Los genes que codifican productos relacionados, aunque estén localizados en diferentes cromosomas, pueden estar ubicados próximos en el núcleo interfásico. Por ejemplo, los cromosomas humanos 13, 14, 15, 21 y 22 poseen un gran número de genes que codifican para ARNr. Dichos cromosomas están agrupados de tal forma que los genes de los ARNr están todos juntos y confinados en el nucléolo, el lugar donde se sintetizan, procesan y ensamblan los ARNr. Esta separación física asegura que los ARNr puedan ser eficientemente ensamblados dentro de las subunidades ribosomales.
En el núcleo, los genes transcripcionalmente activos tienden a estar separados de los inactivos. Los activos se encuentran ubicados centralmente, mientras que los silentes están confinados próximos a la envoltura nuclear.
Tan pronto como las células entran en mitosis o meiosis, los fragmentos de la matriz nuclear dirigen la condensación de los cromosomas, constituyéndose en la parte central de los mismos.


1.2. La envoltura nuclear
La envoltura está formada por dos membranas concéntricas interrumpidas por poros nucleares y por la lámina nuclear.
Las membranas delimitan un espacio de 10 a 50 nm, el espacio o cisterna perinuclear. La membrana externa en contacto con el citoplasma tiene polisomas adheridos, que sintetizan las proteínas que se vuelcan al espacio perinuclear que se continua físicamente con las cavidades del R.E.R.
La envoltura nuclear es un derivado del sistema de endomembranas, siendo esto evidente al inicio de la división celular, cuando la envoltura se desorganiza y pasa a formar parte del sistema de cisternas y vesículas del retículo endoplásmico.
La aparición de la envoltura nuclear permitió que los eucariontes aislaran los procesos genéticos principales, como la autoduplicación del ADN o la síntesis de ARN. Además esto posibilitó que el ARNm se modifique dentro del núcleo antes de ser traducido en los ribosomas. Estas modificaciones no ocurren en los procariontes, ya que a medida que la ARN polimerasa sintetiza el ARN, simultáneamente el extremo 5’ se une al ribosoma y comienza la traducción.
Las membranas nucleares externa e interna se unen, de tanto en tanto, constituyendo en la envoltura perforaciones circulares que poseen siempre el mismo diámetro, son los denominados poros nucleares, gracias a ellos la envoltura no forma una barrera continua entre el nucleoplasma y el hialoplasma.


En el borde de cada poro nuclear se observa la presencia de un material que forma un cilindro, orientado perpendicularmente a la envoltura, que atraviesa el poro y da salida a la vez al nucleoplasma y al hialoplasma, este material constituye el denominado complejo del poro.
Este complejo está constituido por 8 subunidades de ribonucleoproteínas. El complejo del poro disminuye el diámetro del orificio del poro, y además el material del complejo desborda la periferia del poro.
El complejo del poro aparece pues como un anillo cuyo diámetro externo es superior al del poro y cuyo diámetro interno es inferior al del poro, de ahí el nombre de annulus dado al complejo (figura 7.31).


El número de poros presentes en la envoltura nuclear es muy variable dependiendo del tipo celular y la actividad funcional. Si su frecuencia es baja se disponen al azar, sin embargo si su frecuencia es alta adoptan un ordenamiento hexagonal regular. Su función es regular el intercambio de sustancias entre el núcleo y el citoplasma.
A lo largo de la membrana nuclear interna se observa, adosada por fuera de los poros y de sus complejos, una capa cuyo espesor y estructura varía según los tipos celulares y los organismos. Situada entre la membrana nuclear interna y las masas de cromatina periféricas, recibe el nombre de lámina nuclear. Su espesor está comprendido entre 150 y 600 Å, según las células.
La lámina nuclear está constituida por proteínas, a las que se adosan las masas de cromatina. Las proteínas se unen a la membrana interna por medio de otras proteínas de membrana que sirven de anclaje (figura 7.32).

Durante la mitosis la lámina nuclear desaparece, esa desaparición induce la ruptura de la envoltura nuclear. Cuando termina el proceso de división celular colabora en la formación de la nueva envoltura nuclear.
Sus funciones son las siguientes:
a) Dar estabilidad a la envoltura nuclear.
b) Organizar las masas de cromatina.
1.3. Nucleoplasma
También se le conoce como jugo nuclear, carioplasma o cariolinfa. Es una disolución coloidal que constituye el medio interno del núcleo, compuesta por proteínas, ácidos nucleicos, lípidos, glúcidos, sales minerales e iones, disueltos en agua. Sus funciones se reducen a que en su seno tiene lugar la síntesis de ácidos nucleicos.
1.4. Cromatina y cromosomas
El núcleo contiene los cromosomas de la célula. Cada cromosoma consiste en una molécula única de ADN con una cantidad equivalente de proteínas. Colectivamente, el ADN con sus proteínas asociadas se denomina cromatina.
La mayor parte de las proteínas de la cromatina consisten en copias múltiples de cinco clases de histonas. Estas proteínas básicas son ricas en residuos de arginina y lisina cargados positivamente. Por esta razón se unen estrechamente con los grupos fosfatos (cargados negativamente) del ADN.
La cromatina también contiene pequeñas cantidades de una amplia variedad de proteínas no histónica. La mayoría de ellas son factores de transcripción (por ejemplo, el receptor esteroide), siendo su asociación con el ADN pasajera. Estos factores regulan que parte del ADN será transcripta en ARN.
1.4.1 Niveles de organización de la cromatina
La observación a través del microscopio óptico de un núcleo interfásico, nos permite distinguir dos tipos de cromatina. La eucromatina o cromatina laxa o difusa, de localización central, y la heterocromatina o cromatina densa o condensada, en la periferia del núcleo. La heterocromatina representa aproximadamente el 10% del total de cromatina y es considerada transcripcionalmente inactiva.
La heterocromatina puede ser, a su vez, constitutiva, que no se expresa nunca y que sirve de soporte estructural de los cromosomas, o facultativa, que sólo se transcribe en algunos procesos de desarrollo celular como por ejemplo la diferenciación celular, quedando inactivada posteriormente.

La eucromatina se encontraría al menos en dos estados: la eucromatina accesible, que representa alrededor del 10%, donde se encuentran los genes que se están transcribiendo y la eucromatina poco accesible, más condensada (pero menos que la heterocromatina, donde están los genes que la célula no está transcribiendo.
La forma menos condensada de la cromatina tiene la apariencia de un collar de perlas. Las perlas son los nucleosomas, las unidades básicas de enrollamiento de la cromatina. Los nucleosomas están formados por un centro o "core" de histonas. Dicho centro posee dos copias de cada una de las siguientes histonas: H2A; H2B; H3 y H4.
Alrededor del centro de histonas, 146 pares de bases del ADN se enrollan en dos vueltas. La unión de las histonas al ADN no depende de una secuencia particular de nucleótidos, sino de la secuencia de aminoácidos de la histona. Las histonas son unas de las moléculas más conservadas durante el transcurso de la evolución (lo cuál es reflejo de su importancia).
Alrededor de 60 pares de bases de ADN unen un nucleosoma con el próximo. Cada región de unión es el ADN espaciador. La quinta histona, la H1, conecta a los nucleosomas y actúa como una banda de goma, manteniéndolos juntos dentro de una misma cuerda enrollada. Esta estructura se conoce como fibra de 10 nm o collar de cuentas, siendo el primer grado del empaquetamiento de la cromatina.



Recordemos que existe un caso especial de primer nivel de empaquetamiento que resulta de la asociación del ADN con protaminas en lugar de con histonas. Este tipo de asociación aparece en el núcleos de los espermatozoides.
Las protaminas son más pequeñas y más básicas que las histonas, lo que implica una fuerza mayor de atracción entre ellas y el ADN y, consecuentemente, un mayor grado de empaquetamiento del ADN, lo que favorece la movilidad de los espermatozoides. En esta asociación se observa una disposición ordenada de sus componentes, ADN y protaminas, de manera que se ocupa el mínimo espacio posible. Se denomina estructura cristalina (figura 7.35). A diferencia de las histonas, las protaminas son distintas para cada especie.

Los nucleosomas se organizan, a su vez, en fibras de 30 nm o fibra de cromatina de 300 Ǻ. Se forma por enrollamiento, sobre sí misma, de la fibra condensada de cromatina de 100 Ǻ, o sea de la fibra que contiene histonas H1. El modelo más aceptado, recibe el nombre de solenoide, se origina al enrollarse el collar de perlas de tal forma que ese enrollamiento presenta 6 nucleosomas por vuelta (figura 7.36).

En él, las histonas H1 se agrupan formando el eje central de la fibra de 300 Ǻ lo que implica un acortamiento de, aproximadamente, 5 veces la longitud del collar de perlas. En el núcleo celular la mayor parte de la cromatina está en forma de fibra de 100 Ǻ o de 300 Ǻ.


En el siguiente nivel de empaquetamiento, las fibras de 30 nm se organizan en una serie de bucles o asas superenrolladas, tercer grado de empaquetamiento de la cromatina. Estos bucles se estabilizan gracias a la interacción con las proteínas de la matriz nuclear o andamiaje nuclear.
Cada bucle de cromatina representa un dominio funcional o unidad de replicación (Fig. 10.10e). Estos dominios contienen alrededor de 100.000 pares de bases, extensión de ADN suficiente para acomodar varios genes de tamaño promedio. Algunos genes, sin embargo, pueden abarcar varios dominios adyacentes de un cromosoma. Cada cromosoma puede tener cien o más dominios. Durante la profase, los cromosomas aparecen en forma más condensada, alcanzando la cromatina su mayor nivel de condensación en metafase. La organización de los cromosomas envuelve la fosforilación de la H1 y otras proteínas, lo cual causa el plegamiento y empaquetamiento aún más compacto de la cromatina. El andamiaje o matriz nuclear se convierte en el centro de la estructura del cromosoma, y como la compactación continúa, éste se pliega modo de acordeón (cuarto grado de empaquetamiento de la cromatina).


El quinto grado de empaquetamiento de la cromatina ya sería el cromosoma metacéntrico (máxima condensación posible). El grado de condensación de los dominios de cromatina se mantiene principalmente debido a la asociación con la matriz nuclear y a proteínas asociadas como la topoisomerasa II o girasa, encargada de controlar el grado de superenrollamiento del ADN . La unión entre la cromatina y la matriz se da a nivel de zonas altamente conservadas, denominadas secuencias SAR o MAR.


Resumen del empaquetamiento:


1.4.2 El cromosoma eucariota
Fueron observados por primera vez por Hofmeister en 1848, en las células del polen de Tradescantia sp., aunque su nombre se debe a Waldeyer (1888). Su naturaleza nucleica fue descubierta en 1869, aunque hasta 1943 Avery no identificó al ADN como portador de la información genética.
Cada cromosoma eucariota consiste en una molécula simple de ADN de alrededor de 150 millones de pares de nucleótidos.
La molécula de ADN en el cromosoma eucariota es lineal, por lo tanto posee dos extremos (en contraste con el cromosoma bacteriano que es circular).
La molécula de ADN de un cromosoma típico eucariota contiene un conjunto lineal de genes que codifican para ARN y proteínas interrumpido por muchas secuencias de ADN no codificante.
El número de cromosomas de una célula varía según las especies. Existen algunas especies en las que todos sus cromosomas son distintos entre sí, se les llama especies haploides (n). En otras, los cromosomas se pueden agrupar por parejas, de manera que los dos cromosomas de una pareja son idénticos en cuanto a su forma y portan genes para el control de los mismos caracteres, se les llama diploides (2n). En este caso, a los dos cromosomas que forman una pareja se les conoce como cromosomas homólogos.
El mejor momento en el ciclo vital de una célula para estudiar la morfología de los cromosomas es la división celular, y dentro de ella los períodos de metafase y de anafase, que son las dos fases de la mitosis en las que mejor se observan (los cromosomas metafásicos son los mejor estudiados).
Mediante MO, aparecen como bastoncitos de 0,2 a 1 μm de diámetro por 0,2 a 50 μm de longitud, divididos transversalmente en dos brazos por un fuerte estrechamiento, llamado constricción primaria o centrómero, a nivel del cual se une a las fibras del huso acromático durante la división celular. Estos centrómeros están formados por heterocromatina constitutiva (inactiva genéticamente en todas las células).

A ambos lados del centrómero y sobre cada una de las dos cromátidas, se localiza una estructura de naturaleza proteica denominada cinetocoro (lugar desde donde se van polimerizar los microtúbulos que intervienen en la separación de las cromátidas durante la anafase de la mitosis).
Pueden existir constricciones secundarias, relacionadas con la formación del nucleolo al final de la mitosis, por esta razón también reciben el nombre de organizadores nucleolares (NOR).
En los extremos del cromosoma existen unas zonas diferenciales que forman un casquete y que evitan que se pierda información de los extremos en cada ciclo de replicación. Estas zonas reciben el nombre de telómeros. En el ser humano los telómeros contienen la secuencia TTAGGG repetida miles de veces (están formados por heterocromatina constitutiva). Los telómeros presentan las siguientes funciones:
a) Son esenciales para la duplicación del cromosoma.
b) Protegen a los cromosomas frente a las nucleasas (enzimas que digieren el ADN).
c) Evitan que los extremos del cromosoma se fusionen entre sí.
d) Facilitan la interacción entre los extremos y la envoltura nuclear.
En ocasiones, a uno de los extremos se le une un fragmento de ADN denominado satélite, que consiste en un cuerpo más o menos redondeado, no más ancho que el brazo, unido al extremo por un filamento fino.
Cuando los cromosomas se colorean se observa un bandeado de diferente intensidad de coloración que va a permitir una identificación inequívoca de los cromosomas mediante el llamado método de patrón de bandas.
Este método se basa en la utilización de diversas técnicas de coloración tras digestión enzimática o desnaturalización por calor. A estas bandas coloreadas se les ha llamado C, G, Q y R y permiten identificar a los cromosomas homólogos.
También se observan divididos longitudinalmente, formados, por tanto, por dos unidades, las denominadas cromátidas que contienen exactamente la misma información genética ya que una de ellas es copia exacta de la otra, realizándose dicha copia durante el periodo S de la interfase del ciclo celular.
En la anafase, se separan emigrando cada una de ellas a un polo de la célula, por lo que los cromosomas anafásicos de igual morfología que los metafásicos, sólo se diferencian de estos en que poseen una sola cromátida.
Dependiendo de la longitud de los brazos y de la posición del centrómero distinguimos cuatro tipos de cromosomas metacéntricos:
a) Cromosoma metacéntrico, los dos brazos son iguales, el centrómero está en el centro. Cuando se separan las cromátidas en la anafase mitótica adquieren forma de V.
b) Cromosoma submetacéntrico, el centrómero ocupa un aposición submedial. Uno de los brazos tiene un tamaño ligeramente superior al otro. Cuando se separan las cromátidas en la anafase mitótica adquieren forma de L.
c) Cromosoma acrocéntrico, el centrómero ocupa una posición subterminal. Uno de los brazos es muy largo mientras que el otro es muy corto.
d) Cromosoma telocéntrico, el centrómero ocupa uno de los extremos del cromosoma, lo que da lugar a un cromosoma que posee un único brazo.

Otra parte de los cromosomas formada por heterocromatina constitutiva son las múltiples secuencias señalizadoras altamente conservadas, denominadas origen de replicación (ORI), necesarias para que se realice la duplicación del ADN en un tiempo breve.
El cariotipo es una representación gráfica o fotográfica de los cromosomas presentes en el núcleo de una sola célula somática de un individuo ordenados. Cada miembro del par de cromosomas homólogos proviene de cada uno de los padres del individuo cuyas células examinamos. En los seres humanos hay 22 pares de autosomas (pares de cromosomas iguales) y un par de heterosomas o cromosomas sexuales (el 23).
El par 23 de las mujeres tiene dos cromosomas XX y el de los hombres un cromosoma X y un cromosoma Y; en el cromosoma Y hay un gen (el gen SRY) que desencadena la formación de los testículos y, por lo tanto, de los caracteres masculinos.

1.5. El nucleolo
Los nucleolos son estructuras densas y esféricas de número definido en los núcleos interfásicos y profásicos de todos los organismos superiores. Estas estructuras son diferenciaciones cromosómicas funcionales, organizadas en lugares determinados de ciertos cromosomas, concretamente de aquellos que transportan los genes ribosómicos pesados (28S y 18S). En el ser humano, los pares 13,14, 15, 21 y 22, aportan sectores de cromatina que forman el nucléolo. Todos estos cromosomas son acrocéntricos y presentan constricciones secundarias denominadas organizadores nucleolares (NOR), donde están los genes que codifican ARNr. Son el lugar de síntesis de estos ARN-r, de su metabolismo post-transcripcional y de su ensamblaje, por una parte, con los ARN-r ligeros (5S), y por otra, con las proteínas de origen citoplasmático.
Cuando se utiliza MO se observa que en la mayor parte de los sistemas celulares, los nucleolos son formaciones bien individualizadas y fácilmente distinguibles con el microscopio óptico debido a su tamaño (1 a 7 μm de diámetro), a su fuerte refringencia y a su afinidad por los colorantes básicos. Su número en general es de uno o dos por núcleo, si bien existen casos excepcionales en los que puede haber más de diez. Cuando se observan con MET aparecen como una masa, densa a los electrones, de aspecto esponjoso, con cavidades que se encuentran llenas de jugo nuclear o de cromatina (es la cromatina que origina, en la división, el cromosoma organizador del nucleolo). A grandes aumentos se observa formado por un gran número de partículas esferoidales de unos 150 Å de diámetro y fibrillas de 100 Å de sección.
Mediante técnicas citoquímicas y análisis químicos de fracciones nucleolares aisladas, se ha comprobado que en su composición hay: Agua en un 40%, proteínas asociadas al ARN, que forman las fibrillas, fosfolípidos, polisacáridos y ARN, de composición similar al ARN-r, que parece ser organiza las partículas de 150 Å.
En el nucléolo tiene lugar la formación de subunidades ribosómicas, la síntesis y procesamiento de ARNr y actualmente se considera que desempeña un importante papel en la regulación del ciclo celular.
El tamaño del nucleolo varía entre células y en la misma célula según su actividad, pues si bien la velocidad de transcripción puede acelerarse, el ensamblado de las subunidades ribosomales requiere de un tiempo más o menos constante; es por ello que en los nucléolos grandes observamos mayor proporción de componente granular.

2. El ciclo celular
El ciclo celular (también llamado ciclo de división celular) es una secuencia de sucesos que conducen en primer lugar al crecimiento de la célula y posteriormente a la división de la misma en células hijas. Se inicia en el instante en que aparece una nueva célula, descendiente de otra que se ha dividido, y termina en el momento en que dicha célula, por división subsiguiente, origina nuevas células hijas.
El ciclo celular es la base para la reproducción de los organismos. Su función no es solamente originar nuevas células sino asegurar que el proceso se realice en forma debida y con la regulación adecuada (con controles internos para evitar la posible creación de células con múltiples errores). La creación de nuevas células permite al organismo mantenerse en un constante equilibrio, previniendo así aquellos desórdenes que puedan perjudicar su salud (enfermedades congénitas, cáncer, etc.). Los controles internos en la célula son ejecutados por proteínas que no permiten que se presenten situaciones desastrosas (enfermedades) para un ser vivo. Las células que no entrarán en división no se consideran que estén en el ciclo celular.
En rigor, el ciclo celular (la secuencia de sucesos) comprende dos periodos bien nítidos: la interfase (etapas G1, S y G2) y la división celular (etapa M). Esta ultima comprende dos procesos fundamentales: la división nuclear (mitosis o meiosis) y la división del citoplasmas (citocinesis).
2.1 Interfase
La interfase es el período comprendido entre divisiones celulares. Es la fase más larga del ciclo celular, ocupando casi el 95% y, como ya hemos dicho, se divide en tres subetapas: G1, S y G2.
El estado o etapa G1, del inglés Growth o Gap1 (Intervalo 1), es la primera fase del ciclo celular, en la que existe crecimiento celular con síntesis de proteínas y de ARN. Es el período que trascurre entre el fin de una mitosis y el inicio de la síntesis de ADN. Tiene una duración variable (depende del tipo celular), y durante este tiempo la célula duplica su tamaño y masa debido a la continua síntesis de todos sus componentes, como resultado de la expresión de los genes que codifican las proteínas responsables de su fenotipo particular. Al final de esta etapa se distingue un momento de no retorno, llamado punto de restricción o punto R, a partir del cuál ya es imposible detener la sucesión de las fases S, G2 y M.
En este período la célula tiene cromosomas con una sola cromátida y un solo diplosoma (dos centriolos).
Algunas células permanecen en estado de reposo de forma permanente y no se dividen: a esto se le llama período G0
El estado o etapa S (del inglés Synthesis) es la segunda fase del ciclo, en la que se produce la replicación o síntesis del ADN, como resultado cada cromosoma se duplica y queda formado por dos cromátidas idénticas. Con la duplicación del ADN, el núcleo contiene el doble de proteínas nucleares y de ADN que al principio. Tiene una duración de unos 6-8 horas.
El estado o etapa G2 del inglés Growth o Gap2 (Intervalo 2), es el tiempo que transcurre entre la fase S y el inicio de la mitosis (la célula se prepara para mitosis). Tiene una duración entre 3 y 4 horas. Termina cuando la cromatina empieza a condensarse al inicio de la mitosis y los cromosomas se hacen visibles.


El final de la mitosis da cabida a un nuevo ciclo en G1 o puede que la célula entre en fase G0 que corresponde a un estado de reposo especial característico de algunas células, en el cual puede permanecer por días, meses y a veces años. Las células que se encuentran en el ciclo celular se denominan proliferantes y las que se encuentran en fase G0 se llaman células quiescentes.
Como todo proceso orgánico, el ciclo celular está sujeto a regulación. Ésta es realizada en sitios específicos llamados puntos de control o de chequeo, que pueden frenar o disparar diversos procesos que le permitan a la célula proseguir con su ciclo normal de replicación del material genético, crecimiento y división. La función de la regulación, básicamente es realizada por proteínas específicas conocidas como cinasas (kdc) y ciclinas (ciclinas A ó B).
Hay células que se encuentran permanentemente en el ciclo, como las epiteliales; otras están permanentemente fuera del ciclo, como las neuronas, y otras están fuera del ciclo, pero bajo un estímulo adecuado pueden volver a dividirse, como es el caso de las células hepáticas.
Durante la fase anterior G1, las moléculas de ADN están enrolladas alrededor de las proteínas histonas, formando los fibras de cromatina que constituyen los cromosomas. Durante la fase S se realiza la duplicación de los cromosomas en el núcleo, por replicación de las moléculas de ADN. Las dos moléculas hijas de cada cromosoma son idénticas entre sí, por lo que se denominan cromátidas hermanas.
Durante esta fase se inicia la condensación de los cromátidas hermanas hasta formar los cromosomas mitóticos. Cada cromátida tiene forma de bastoncillo y permanecen unidas entre si a través de una región denominada centrómero.
La mitosis es la fase en la cual los núcleos de las células se dividen y se reparten las cromátidas hermanas. Es un proceso continuo en el que se distinguen cinco etapas: Profase, prometafase, metafase, anafase y telofase.
3.1 Profase
A lo largo de esta fase finaliza la condensación de las cromátidas hermanas iniciada en la fase G2. Los cromosomas mitóticos se van haciendo más cortos y gruesos, haciéndose visibles al microscopio óptico.
El complejo centriolar (formado por dos centriolos y fibras pericentriolares), se duplica formando los ásteres que comienzan a alejarse a polos opuestos del núcleo, al tiempo que se forman nuevas fibras de microtúbulos polares que constituyen el huso mitótico o acromático.
Las células animales, que poseen centriolos y por tanto forman aster realizan una mitosis astral, para diferenciarla de la mitosis anastral que realizan las células vegetales que carecen de centriolos. En este caso las fibras del huso se forman en una región del citoplasma cercana al núcleo y libre de orgánulos que se conoce como zona clara.
El nucleolo se va fragmentando hasta que desaparece.


La ruptura de la envoltura nuclear señala el comienzo de esta fase, quedando los cromosomas libres.
Se forman los microtúbulos cinetocóricos, a partir de los cinetocoros de los cromosomas. Estos microtúbulos son perpendiculares al eje del cromosoma.
Al final de esta fase los cromosomas comienzan a orientarse respecto a los polos, de modo que cada cinetocoro se encuentra frente a un de los polos.

Los cromosomas se disponen con sus cinetocoros en un mismo plano ecuatorial equidistantes de los dos polos, formando la llamada placa metafásica o ecuatorial. Los microtúbulos cinetocóricos se alargan haca los polos.
Los cromosomas están condensados al máximo (cromosomas metacéntricos).


3.4 Anafase
Las cromátidas hermanas de cada cromosoma se separan y se convierten en cromosomas independientes que son desplazados, a la misma velocidad y simultáneamente, hacia el polo que miran sus cinetocoros (son los que arrastran al resto del cromosoma).
Esto es posible por un acortamiento de los microtúbulos cinetocóricos y un alargamiento de los microtubulos polares que alargan el huso y alejan ambos polos.


3.5 Telofase
Desaparecen las microtúbulos cinetocóricos y fragmentos de retículo endoplasmático se unen a los cromosomas, formando un esbozo de la envoltura nuclear.
Los microtúbulos polares se agrupan en haces y se alejan de los polos.
Al final de esta fase se completa la envoltura nuclear, los cromosomas comienzan a descondensarse y los microtúbulos polares se encuentran agrupados formando un haz único. El nucleolo se va a empezar a formar junto al cromosoma en el que estaba cuando desapareció.


3.6 Citocinesis
La mitosis, en sentido estricto, es la división del núcleo celular pero también debe producirse la división del citoplasma y la separación en dos, o más, células hijas: esto es la citocinesis. Debido a la existencia de la pared celular el proceso es diferente en las células animales y las células vegetales:

3.6.1 Citocinesis en las células animales
Al final de la anafase, la membrana citoplasmática se invagina, gracias al anillo contráctil de fibras de actina dispuesto alrededor de la célula, iniciando así la formación del surco de división.
A lo largo de la telofase el surco crece y se comprime alrededor del haz de microtúbulos polares.
Al final de la telofase las células hijas solo están unidas por un puente citoplasmático donde se encuentra el haz de microtúbulos.
Por último, el puente citoplasmático se rompe, las células se separan y una de ellas arrastra los restos del haz de microtúbulos que finalmente serán degradados.
3.6.2 Citocinesis en las células vegetales
Durante la telofase, vesículas provenientes del aparato de Golgi confluyen en la región media de la célula y se fusionan formando el fragmoplasto.
Al final de la telofase todas las vesículas se han fusionado formando las membranas plasmáticas de las dos células hijas. A partir del contenido de la vesículas se forma la pared celular (lámina media). Los puentes o poros citoplasmáticos persisten entre las dos células hijas y constituyen los plasmodesmos.

La división mitótica es conservativa, es decir las células hijas presentan exactamente el mismo número y el mismo tipo de cromosomas que la célula madre. Pueden llevarla a cabo tanto células diploides (2n) como haploides (n).

Sus objetivos son:
a) En los organismos unicelulares con reproducción asexual la formación de nuevos individuos.
b) En los organismos pluricelulares originar nuevas células para el crecimiento y desarrollo del individuo, sustituir a las células muertas por el desgaste normal, regenerar las partes del organismo destruidas o perdidas y en muchos casos producir células especiales para la reproducción.
Esta división nuclear es especial y su función es formar los gametos (imprescindibles para llevar a cabo la reproducción sexual). Los gametos son células haploides (n) con una sola copia de cada cromosoma mientras el resto de nuestras células son diploides (2n) con dos copias de cada cromosoma. Por eso a la meiosis muchas veces se le denomina división reduccional.
La meiosis va a consistir en dos divisiones consecutivas: una primera división meiótica en la que van a separarse los pares de cromosomas y una segunda división meiótica donde van a separarse las dos cromátidas hermanas de cada cromosoma (de forma similar a como ocurría en la mitosis).
4.1 Primera división meiótica
La dividimos en cuatro fases: profase I, metafase I, anafase I y telofase I (el I se indica siempre en numeración romana y no latina).
4.1.1 Profase I
Es la fase clave de la meiosis y la más larga y compleja. Durante todo el proceso la envoltura nuclear permanece intacta, al tiempo que se desintegra el nucleolo y se forma el huso mitótico. Se distinguen cinco fases:
a) Leptoteno; los cromosomas se hacen visibles por condensación. Sin embargo no se distinguen las cromátidas hermanas pues permanecen estrechamente unidas. Los cromosomas forman bucles de manera que sus extremos se unen a la cara interna de la envoltura nuclear a través de la placa de unión.


b) Zigoteno; los cromosomas homólogos se aparean punto por punto, de forma intima, en toda su longitud. Este emparejamiento gen a gen de los cromosomas homólogos se denomina sinapsis. Cada par cromosómico resultante de la sinapsis se denomina cromosoma bivalente o tétrada.
El apareamiento preciso de los homólogos queda estabilizado y mantenido hasta el final de la siguiente fase gracias a la formación de una estructura proteica en forma de cinta denominada complejo sinaptonémico.

La sinapsis se produce entre todos los cromosomas homólogos excepto entre los cromosomas X e Y, que solo se aparean parcialmente.

c) Paquiteno; los cromosomas se encogen haciéndose más gruesos. Durante esta fase se produce el intercambio de fragmentos cromatídicos entre cromátidas no hermanas de los cromosomas homólogos, mediante un procesos complejo conocido como entrecruzamiento o sobrecruzamiento y que es la causa de la recombinación genética. Este proceso es muy importante pues aumenta enormemente la variabilidad genética.
Los puntos de sobrecruzamiento corresponden a los nódulos de recombinación de los complejos sinaptonémicos formados en la fase anterior,
que contienen las enzimas necesarias para el intercambio de genes entre las cromátidas de los dos cromosomas homólogos (endonucleasas, ADN-polimerasa, ADN-ligasa y otras proteínas imprescindibles para que se produzca el sobrecruzamiento). Este proceso implica la rotura de la doble hélice en los fragmentos que van a ser intercambiados y una posterior fusión en su nueva localización (figura 8.6).

Las cromátidas hermanas ya son claramente visibles, de manera que cada cromosoma bivalente se observa claramente como una tétrada.

d) Diploteno; desaparece el complejo sinaptonémico lo que provoca la separación de los cromosomas homólogos. La separación no es total pues las cromátidas no hermanas permanecen unidas por zonas denominadas quiasmas, que son los puntos por donde se produjo el entrecruzamiento. Esta etapa es la más larga de la meiosis, puede durar días e incluso años (como es el caso de los ovocitos humanos).



e) Diacinesis; la condensación de las tétradas es máxima. Las cromátidas no hermanas permanecen unidas por los quiasmas que se han desplazado a los extremos de los cromosomas homólogos. Al final de esta fase comienza la desintegración de la envoltura nuclear y del nucleolo, al tiempo que se duplica el nucleosoma, se forma el aster y se inicia la formación del huso con microtúbulos polares.


4.1.2 Metafase I
Se forman los microtúbulos cinetocóricos, pero a diferencia de la mitosis, los dos cinetocoros de un mismo cromosoma homólogo están orientados hacia el mismo polo, mientras que los cinotocoros del homólogo se orientan hacia el polo opuesto.
Esto ocurre porque se van a separar los cromosomas homólogos y no las cromátidas. Es muy importante no confundirlo (y el dibujo) con la metafase de la mitosis.

4.1.3 Anafase I
Los quiasmas se rompen y, al acortarse los microtúbulos cinetocóricos, se produce la separación de los cromosomas homólogos, que migran hacia polos opuestos.

4.1.4 Telofase I
Desaparece el huso y se forma la envoltura nuclear alrededor de los dos núcleos hijos. Los cromosomas se sufren una ligera descondensación y la célula se divide en dos (citocinesis).

Como resultado de esta primera división meiótica se producen dos células hijas que tienen cada una de ellas un juego completo de cromosomas homólogos procedentes de la célula madre (haploides). Estos cromosomas homólogos ya no son completamente paternos o maternos pues debido al entrecruzamiento alternan algunos segmentos paternos y maternos al azar. De esta forma la recombinación genética hace que todos los gametos sean genéticamente diferentes.


4.2 Segunda división meiótica
Una vez terminada la primera división meiótica se produce un breve periodo de interfase (sin síntesis de ADN) en el que los cromosomas se descondensan un poco pero enseguida vuelven a condensarse y se desencadena la segunda división meiótica (muy similar a la mitosis que ya hemos estudiado).
Vamos a distinguir cuatro fases principales: profase II, metafase II, anafase II y telofase II; seguidas por la división del citoplasma o citocinesis.
a) Profase II; es una fase muy breve pues los cromosomas ya estaban muy condensados. Se rompe la envoltura nuclear y se forma el huso mitótico.

b) Metafase II; los cromosomas se alinean en la placa metafásica disponiéndose de tal manera que cada cromátida hermana está enfocada a uno de los polos.

c) Anafase II; se separan las cromátidas hermanas y comienzan a migrar hacia los polos.

d) Telofase II; se forman las envolturas nucleares alrededor de los cuatro núcleos haploides.

Al final se produce la citocinesis ocasionando las cuatro células hijas haploides correspondientes.


Sobre un plano cuantitativo el núcleo diploide (2n) del comienzo de la meiosis se divide durante la primera división meiótica en dos núcleos haploides (n), en los que cada cromosoma está formado por dos cromátidas.
Durante la segunda división meiótica, se producirá una separación de esas cromátidas, es decir esta segunda división meiótica es exactamente igual a una mitosis. Al final del proceso la célula diploide (2n) original que se ha dividido ha dado lugar a cuatro células haploides (n).
Lo más característico del proceso es que se ha producido un intercambio, durante la profase I, de fragmentos entre cromosomas homólogos, que tiene como consecuencia una recombinación genética, lo que, a su vez, es causa de la alta variabilidad genética que presentan los individuos de una misma especie.
La meiosis es un proceso imprescindible para los organismos que se reproducen mediante reproducción sexual, es decir aquellos que requieren de la unión de dos células, denominadas gametos, cada una de las cuales procede de un progenitor distinto.
El proceso de fusión de los gametos recibe el nombre de fecundación y tiene como resultado la formación de una célula denominada cigoto que es la primera célula del nuevo individuo.
Si los gametos fueran diploides, al unirse el cigoto sería tetraploide (4n), con lo que el nuevo individuo no tendría la misma dotación genética que sus progenitores. Para evitar esto se ha desarrollado el mecanismo de meiosis que produce gametos haploides (n), cuando estos gametos se unen el nuevo individuo vuelve a ser diploide (2n) como sus progenitores.
La meiosis implica tres efectos importantes:
a) Reducción del número de cromosomas durante la formación de los gametos.
b) Modificación de la información genética de los cromosomas durante el sobrecruzamiento de la profase I.
c) Distribución de los cromosomas entre los gametos, lo que permite que durante la fecundación se mezclen al azar.
La consecuencia de tales efectos es de un lado el mantenimiento de la constancia genética de las especies y de otro el enorme potencial de diversidad genética de las especies que se reproducen sexualmente, la denominada variabilidad genética. Esto es lo que diferencia a los organismos que se reproducen sexualmente de los que lo hacen asexualmente, en estos últimos la única fuente de variabilidad son las mutaciones.
Podemos concluir que la reproducción sexual, con sus dos procesos básicos de meiosis y de fecundación, potencia enormemente la variabilidad genética, lo que es la base del proceso evolutivo.
5. Principales diferencias entre la mitosis y la meiosis



No hay comentarios:
Publicar un comentario