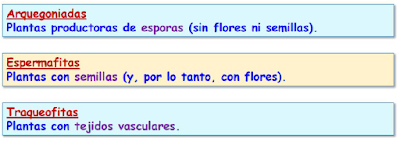
Las plantas son seres vivos eucariotas, pluricelulares, fotosintéticos, que han colonizado el medio terrestre gracias a la aparición de un tejido, la epidermis, que aísla de la desecación al individuo. También han desarrollado estructuras para fijarse al sustrato y absorber agua y sales minerales. Evolutivamente proceden de las clorofitas o algas verdes.

La reproducción puede ser asexual o sexual. La reproducción sexual se realiza mediante la unión de células gaméticas de distinto tamaño. El gameto masculino se denomina genéricamente anterozoide y el gameto femenino, oocito u ovocito. El zigoto, formado al unirse los gametos, origina un embrión pluricelular.
Todas las plantas tienen un ciclo biológico diplohaplonte, con alternancia de una fase haploide, denominada gametofito que produce gametos masculinos y femeninos, y otra diploide. La fase diploide se produce cuando, por la fecundación de los gametos, se forma el cigoto que origina el esporofito. En el esporofito se produce la meiosis, originando esporas que generan de nuevo el gametofito.
Evolutivamente, la fase gametofítica disminuye, llegando a ser microscópica en las plantas más evolucionadas. Por el contrario, el esporofito pasa, de ser una estructura que sólo aparece en épocas reproductivas en plantas menos evolucionadas, a ser una estructura macroscópica, con crecimiento anual, en plantas superiores.




1. La célula vegetal

2. Histología vegetal
Cuando hablamos de las características de los tejidos de las plantas tenemos que tener en mente la historia ocurrida hace 500 millones de años, cuando las plantas conquistaron la tierra. El medio terrestre ofrece ventajas respecto al medio acuático: más horas y más intensidad de luz, y mayor circulación libre de CO2.
Pero a cambio las plantas tienen que solventar nuevas dificultades, casi todas relacionadas con la obtención y retención de agua, con el mantenimiento de un porte erguido en el aire y también con la dispersión de las semillas en medios aéreos. Para ello las plantas agrupan sus células y las especializan para formar tejidos con funciones determinadas que sean capaces de hacer frente a estas nuevas dificultades.


Para superar un medio ambiente variable y seco, aparece un sistema protector formado por dos tejidos: la epidermis y la peridermis. Las células de estos tejidos se revisten de cutina y suberina para disminuir la pérdida de agua, y aparecen los estomas en la epidermis para controlar la transpiración y regular el intercambio gaseoso.
Para mantenerse erguidas sobre la tierra las plantas tienen un sistema de sostén representado por dos tejidos: el colénquima y otro más especializado denominado esclerénquima.
Una gran cantidad del tejido de las plantas es el parénquima, el cual realizará diversas funciones, desde la fotosíntesis hasta el almacén de sustancias.
Uno de los hechos más relevantes en la evolución de las plantas terrestres es la aparición de un sistema conductor capaz de comunicar todos los órganos del cuerpo de la planta, formado por dos tejidos: el xilema, que conduce mayormente agua, y el floema, que conduce principalmente sustancias orgánicas en solución. Sólo hablamos de verdaderos tejidos conductores en las plantas vasculares.


Finalmente, las plantas vasculares producen semillas, dentro de las cuales se forma el embrión, que se desarrolla y crece gracias a la actividad de los tejidos embrionarios o meristemáticos.
Los meristemos, no sólo están presentes en el embrión sino que están activos a lo largo de toda la vida de la planta, permitiendo su crecimiento. Todos estos tejidos, excepto los meristemos, han derivado a lo largo de la evolución de otro tejido poco diferenciado llamado parénquima, que se mantiene en las plantas actuales y que realiza múltiples funciones.

2.1. Meristemos o tejidos vegetales embrionarios
Estos tejidos son los responsables del crecimiento longitudinal y transversal de las plantas y están presentes durante toda la vida de éstas.
Están constituidos por células indiferenciadas, de pequeño tamaño, isodiamétricas y tienen una pared celular primaria delgada, careciendo de pared secundaria, ya que van a modificar su estructura durante su diferenciación.
Su citoplasma contiene abundantes ribosomas, un retículo endoplasmático rugoso escaso, el complejo de Golgi muy desarrollado para fabricar los componentes de la pared celular, numerosos plastos en desarrollo (proplastidios), muchas y pequeñas vacuolas y un protoplasma desprovisto de inclusiones. El núcleo, con mucha cromatina condensada, es grande y se sitúa en posición central.

Las células meristemáticas son células totipotentes, están continuamente dividiéndose por mitosis y posteriormente se diferencian para originar el espectro entero de tipos celulares de una planta adulta. La clasificación de los meristemos se realiza en base a su posición en el cuerpo de la planta.
Hay dos tipos de meristemos: primarios (crecimiento longitudinal) y secundarios (crecimiento transversal).

2.1.1. Meristemos primarios
Son los responsables del crecimiento longitudinal de la planta. Entre ellos están:
A) Meristemos apicales: Se sitúan en el ápice del tallo y de cada rama, y en el ápice de la raíz principal y de las raíces secundarias. En el primer caso están protegidos por los primordios foliares, mientras que en el segundo por la cofia o caliptra. Estos meristemos se denominan meristemos apicales caulinares y meristemos apicales radicales, respectivamente.



En cada meristemo apical nos encontramos células que forman la protodermis, que origina la epidermis, el procambium, que produce los tejidos conductores primarios (xilema y floema primarios), y el meristemo fundamental, que dará lugar al resto de los tejidos de la planta.

B) Meristemos intercalares: Se encuentran intercalados entre tejidos no meristemáticos. Se localizan en los entrenudos o en la vaina de las monocotiledóneas.

2.1.2. Meristemos secundarios o laterales
Éstos son responsables del crecimiento en espesor por lo que sus células se dividen en paralelo a la superficie del tallo o de la raíz. Estos meristemos no existen en muchas plantas herbáceas, como la mayoría de las pteridofitas y monocotiledóneas, ni en determinados órganos como las hojas.
Hay dos tipos de meristemos secundarios:
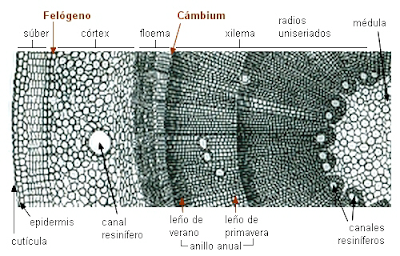
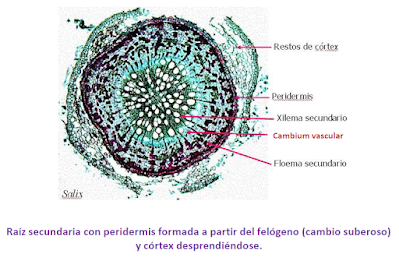
a) Cambium vascular: los cambiums de todos los haces vasculares se conectan entre sí formando un cilindro vascular (anillo continuo) alrededor de planta, produciendo más xilema secundario hacia el interior del tallo y floema secundario hacia afuera. De este modo el tallo presenta los tejidos vasculares formando círculos concéntricos. Las partes vivas del tallo leñoso son las más próximas al cambium.





El cambium produce elementos de mayor diámetro en primavera (leño temprano) y de menor diámetro y paredes más gruesas en invierno (leño tardío); la actividad de todo el año forma así un anillo de crecimiento. Cada año se suma un nuevo anillo de crecimiento, los cuales pueden contarse a simple vista en el corte transversal de un tronco. La presencia de radios medulares, que corren horizontalmente, desde el centro del tallo a la periferia, además de los anillos de crecimiento son las principales diferencias entre los tejidos vasculares del cuerpo primario de la planta y el xilema y floema formados por el cambium.

Albura y duramen: para analizar las características anatómicas de la madera, generalmente se observa al microscopio la parte correspondiente al leño; denominándose como tal al xilema secundario originado a partir del cambium, meristemo lateral o secundario.
En la mayoría de los árboles la parte interna del leño cesa su actividad conductora y sus células vivas (parenquimáticas) mueren, debido fundamentalmente a ciertos cambios:
Desintegración del protoplasma.
Reforzamiento de las paredes con más lignina.
Acumulación en el lumen o impregnación de las paredes con sustancias orgánicas e inorgánicas: taninos, aceites, gomas, resinas, colorantes, compuestos aromáticos, carbonato de calcio, silicio.
Bloqueo de vasos conductores.
El leño que ha sufrido estos cambios es el duramen, inactivo y más oscuro. Cuanto mayor es la impregnación, mayor es la resistencia a los microorganismos que provocan la putrefacción. La porción clara, externa, activa, con células vivas es la albura. La proporción albura-duramen varía en las distintas especies, como también varía el grado de diferenciación entre ambas.

b) Cambium suberoso o felógeno, que origina la peridermis o corteza. Ambos meristemos se disponen, como un cilindro continuo o como un anillo incompleto formando bandas, entre el xilema secundario y el floema secundario para el caso del cambium vascular o entre la felodermis y el súber para el cambium suberoso o felógeno.


2.2. Tejidos vegetales definitivos
Las plantas tienen tres tipos básicos de tejidos:
a) El tejido fundamental comprende la parte principal del cuerpo de la planta. Las células parenquimáticas (las más abundantes), colenquimáticas y esclerenquimáticas constituyen los tejidos fundamentales.
b) El tejido epidérmico cubre las superficies externas de las plantas herbáceas, está compuesto por células epidérmicas fuertemente unidas que secretan una capa formada por cutina y ceras llamada cutícula que impide la pérdida de agua. En él se pueden observar estomas, tricomas y otro tipo de especializaciones.
c) El tejido vascular está compuesto por dos tejidos conductores: el xilema y el floema, transportan nutrientes, agua, hormonas y minerales dentro de la planta. El tejido vascular es complejo, incluye células del xilema, floema, parénquima, esclerénquima y se origina a partir del cambium.

2.2.1. Parénquima
Es un tejido simple y poco especializado, formado por células vivas en la madurez y que conservan su capacidad de dividirse.




Cumplen diversas funciones, de acuerdo a la posición que ocupan en la planta, presentando formas y contenidos celulares acordes:
a) Tejido parenquimático fundamental; es el menos especializado, son células isodiamétricas, de paredes primarias delgadas; se encuentra como relleno entre otros tejidos, en la región medular y en el córtex. Retiene su capacidad de dividirse por mitosis a la madurez, esta característica permite que de una sola célula se pueda regenerar una planta completa por cultivo in vitro.
b) Tejido parenquimático clorofílico o clorénquima; está especializado en la fotosíntesis gracias a que sus células contienen numerosos cloroplastos. Se encuentra por lo general debajo de la epidermis, donde la luz llega más fácilmente, sobre todo en las hojas, aunque también es común en la zona superficial (córtex) de los tallos verdes.
El clorénquima de la hoja se denomina mesófilo y se divide en dos tipos: en empalizada, más expuesto al Sol, y parénquima lagunar, en la parte más sombría. El primero tiene mayor número de cloroplastos y parece llevar a cabo una mayor tasa de fotosíntesis, estando sus células además más densamente empaquetadas. En el parénquima lagunar hay más espacios intercelulares gracias a los cuales es un buen tejido para el intercambio de gases y agua con la atmósfera.








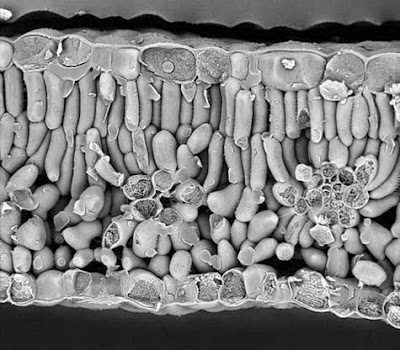
c) Tejido parenquimático de reserva;
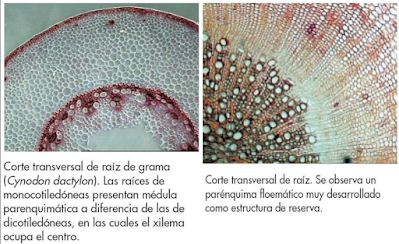
Sus células sintetizan y almacenan diversas sustancias como azúcares en diversas formas, cristales proteicos, proteínas, lípidos, pigmentos, etc. Algunas de estas sustancias pueden encontrarse en forma sólida, aunque lo normal es que estén disueltas en la vacuola, que es el orgánulo especializado en el almacén de sustancias. También en el citoplasma y en los plastidios se pueden acumular algunas sustancias como azúcares y sustancias nitrogenadas. Algunas células almacenan un solo tipo de sustancia aunque otras pueden contener una mezcla de sustancias.
El producto de reserva más frecuente en los tejidos vegetativos (que no son semilla ni fruto) son los carbohidratos, y los almacenan de dos formas: en forma de almidón y en forma de sucrosa o sus derivados (sobre todo fructanos).
El almidón se almacena en los amiloplastos mientras que los derivados de la sucrosa se acumulan en la vacuola.
Las proteínas almacenadas suelen ser importantes como fuente de nitrógeno, un bien escaso para la célula, y generalmente el destino de estas proteínas es la degradación. Proteínas y lípidos se almacenan en el parénquima de muchas semillas, en plastidios denominados proteinoplastos y elaioplastos, respectivamente.
En algunas zonas de las plantas el parénquima puede almacenar taninos y antocianinas. El color de las flores se debe a que algunas células almacenan pigmentos en los cromoplastos o en las vacuolas.
La distribución en la planta del tejido parénquimático de reserva es diversa, y puede encontrarse en la raíz, tallo, hojas y frutas. Por ejemplo, la caña de azúcar y la patata almacenan material de reserva en el parénquima del tallo, y la zanahoria lo hace en el de la raíz. Otro lugar de almacenamiento es el parénquima que forma parte de los radios parenquimáticos horizontales de los haces vasculares, importantes para la reserva de los troncos en el invierno.






d) Tejido parenquimático acuífero; aunque todas las células parenquimáticas almacenan agua en mayor o menos medida, las células del parénquima acuífero están especializadas en esta función.
Las células parenquimáticas que almacenan agua son grandes, de paredes delgadas y con una gran vacuola donde se acumula el agua. En el citoplasma o en la vacuola hay mucílagos (conjunto de sustancias que aumentan la capacidad de absorción y retención de agua).
Este parénquima es característico de las plantas que viven en climas secos (plantas xerófitas). En los órganos subterráneos encargados de almacenar sustancias de reserva no suele haber tejidos especializados en el almacén de agua, aunque las células que contienen almidón u otras sustancias de reserva tienen también una gran capacidad de almacenar agua.


e) Tejido parenquimático aerífero o aerénquima; es un tejido que contiene grandes espacios intercelulares vacíos, mayores que los normalmente encontrados en otros tejidos, por donde circulan los gases que permiten la aireación de los órganos de la planta.

Este parénquima está especialmente desarrollado en las plantas que viven en ambientes muy húmedos o acuáticos (plantas hidrófitas), aunque también puede aparecer en algunas especies no acuáticas sometidas a estrés. Aparece tanto en raíces como en tallos.
El aerénquima es continuo desde los tallos hasta las raíces y los grandes espacios intercelulares permiten la conducción de gases, aumentando la difusión de éstos desde las hojas hasta las raíces. Esta comunicación permite a las plantas que viven en suelos húmedos o anegados mantener un nivel de oxígeno suficiente para la respiración. En suelos encharcados también es un vehículo para la liberación a la atmósfera de gases presentes en las raíces, como el etileno. Este parénquima puede considerarse como una adaptación de las plantas a la hipoxia de suelos anegados.
Las plantas con aerénquima se consideran como uno de los vehículos importantes para el paso de gases de efecto invernadero como el metano desde el suelo, pasando por la raíz y el tallo, hasta la atmósfera. Esto es particularmente importante en cultivos extensivos como los de arroz.



2.2.2. Tejidos vegetales de sostén (colénquima y esclerénquima)
Estos tejidos se encargan de proporcionar consistencia y rigidez a la planta. Se encuentran, por tanto, en zonas que se encargan de esta función, como el tallo y la raíz. Para llevar a cabo su función, sus células presentan paredes celulares muy engrosadas y, en ocasiones, aparecen reforzadas por lignina. Existen dos tipos de tejidos de sostén: el colénquima y el esclerénquima.

2.2.2.1. Colénquima
Las paredes celulósicas de sus células están engrosadas, pero no están lignificadas. Son células vivas, resistentes y extensibles. Su función principal es servir como tejido de sostén a los órganos en crecimiento (tallos herbáceos, hojas, flores) y está ausente en las plantas monocotiledóneas.
No aparece en las raíces ni en estructuras con crecimiento secundario (donde se desarrolla el esclerénquima) y se sitúa bajo la epidermis o el parénquima en forma de cilindro o de bandas discontinuas. Con el tiempo, sus células se endurecen y esas partes de la planta que ya no van a crecer más se tornan duras y frágiles. El colénquima se localiza debajo de la epidermis de los tallos herbáceos y en las ramas jóvenes de los árboles.
Podemos destacar los siguientes tipos:
a) Colénquima angular; en este tipo de colénquima los espesamientos de la pared celular se producen en el ángulo donde convergen varias células, de ahí la ausencia de espacios intercelulares. Este tipo de engrosamientos se disponen a modo de vigas que se orientan longitudinalmente en el órgano, que le confiere una gran resistencia mecánica.

b) Colénquima laminar; en este tipo de colénquima los engrosamientos de la pared celular primaria se manifiestan en las paredes externa e interna, es decir, en las paredes paralelas a la superficie del órgano. En este caso se encuentra en un tallo con crecimiento secundario.



c) Colénquima lagunar; se parece al colénquima angular ya que los engrosamientos de las paredes celulares primarias son mayores en el lugar donde confluyen tres células. Sin embargo, dichos engrosamientos no obliteran completamente el espacio intercelular por lo que se pueden observar espacios vacíos entre las paredes celulares de las células contiguas.




d) Colénquima anular; este tipo de colénquima se distingue del resto por ser el único en donde los espesamientos son uniformes alrededor de la célula. Esto hace que este tipo de colénquima presente espacios intercelulares.

2.2.2.2. Esclerénquima
Es un tejido que se encuentra en órganos y tejidos adultos, es decir, en tejidos y órganos que han terminado su crecimiento en longitud.
Proporciona dureza y rigidez a la planta. Protegen las partes más vulnerables a estiramientos, pesos, presiones y flexiones (por eso son más abundantes en los tallos y ramas).
Sus células son duras, con una pequeña cavidad llamada lumen y están muertas con paredes secundarias lignificadas y/o mineralizadas.
Se diferencian dos tipos de células en este tipo de tejido:
a) Esclereidas o células pétreas; son células poligonales, pequeñas y muy lignificadas, que se disponen aisladas y distribuidas al azar en algunos frutos o bien formando gruesas capas sólidas formando los huesos de los frutos. Con formas estrelladas, presentan una pared celular secundaria extremadamente gruesa que dejan un interior celular muy angosto.


b) Fibras; son células alargadas y con extremos puntiagudos, que pueden presentarse dispersas entre el parénquima o formando grupos en raíces, tallos, hojas y frutos.
El grosor de su pared celular 2ª y su lignificación depende de las hormonas vegetales pero en su madurez suelen estar muertas y todo el espacio del protoplasma celular está ocupado por la pared celular.
La clasificación de las fibras de esclerénquima se basa en su localización.
En el tallo cuando las fibras se encuentran debajo de la epidermis se denominan fibras corticales. Protegiendo al haz vascular están las fibras perivasculares. También formando parte del xilema primario se pueden observar fibras denominadas xilares.

Debido a su alta resistencia a la tensión son de gran importancia económica en la manufactura de tejidos (industria textil).

Los tejidos de protección forman la parte más externa de los órganos de las plantas y se encuentran en contacto con el medio ambiente. Los tejidos de protección típicos son la epidermis y la peridermis, dependiendo de si la planta tiene crecimiento primario o secundario, respectivamente.


2.2.3.1. La epidermis
Es la capa superficial de células que revisten todas las partes del cuerpo primario de la planta, es decir, es la capa de células más externa del cuerpo primario de la planta. Las células de tejido epidérmico se caracterizan por estar vivas, son de diferentes formas, de paredes no muy engrosadas, sin espacios intercelulares, con protoplasma parietal con gran cantidad de agua, con vacuolas grandes, generalmente sin cloroplastos (pero con leucoplastos) y con núcleo parietal.

Las funciones de la epidermis son:
a) Proporciona mayor resistencia mecánica.
b) Protege a los tejidos subyacentes de la sequedad.
c) Controla el intercambio de gases y la pérdida de vapor de agua.
La pared celular primaria de las células epidérmicas está recubierta externamente por una cutícula que disminuye la pérdida de agua y que está formada mayormente por cutina y por ceras, secretadas por la propia célula. Hay epidermis que no poseen cutícula, como se observa en las zonas de absorción de las raíces. En otros casos son muy finas, de grosor intermedio o gruesas y muy gruesas. En este último caso las células epidérmicas muestran pared celular secundaria suberificada.




En la epidermis se encuentran los estomas. Las células oclusivas de los estomas son células epidérmicas especializadas que se organizan para dejar un poro u ostiolo entre ellas a través del cual se pone en contacto el medio interno de la planta con el exterior. Existe una cámara de aire bajo el ostiolo denominada cámara subestomática. Ambas estructuras, junto con las células oclusivas, forman lo que típicamente se denomina complejo estomático. Las células oclusivas tienen forma arriñonada o de palillo de tambor, presentan cloroplastos y una pared celular engrosada de manera no uniforme que posibilita que los cambios de turgencia puedan variar su morfología y de este modo aumentar o disminuir el diámetro del ostiolo.






Los tricomas o pelos también son células epidérmicas especializadas y pueden ser de protección o glandulares, y a veces se usan como carácter taxonómico, es decir, sirven para clasificar especies.







Funcionalmente, los tricomas son prolongaciones epidérmicas que realizan numerosas funciones como evitar herbívoros, guiar a los polinizadores, controlar la temperatura y desecación de las hojas, o proteger frente a un exceso de luz.
En la epidermis de la raíz están los denominados pelos radicales, los cuales sirven para absorber agua y sales minerales. Los pelos radicales son células epidérmicas modificadas que crecen a modo de columna perpendicularmente a la superficie de la raíz. En ellos se encuentran también numerosos microorganismos simbiontes tales como las bacterias fijadoras de nitrógeno. Los pelos radicales aparecen en la zona de maduración de la raíz y se diferencian a partir de células epidérmicas indiferenciadas denominadas tricoblastos. Su patrón (número y distribución) es característico de cada especie, aunque también depende de las condiciones del suelo.

Las células buliformes son células epidérmicas especializadas encargadas de almacenar agua, interviniendo en la marchitez y despliegue de las hojas por transpiración.

2.2.3.2. La peridermis o corteza
La peridermis se desarrolla en las plantas de crecimiento secundario por la actividad del cambium suberoso o felógeno. Se origina por la desdiferenciación del parénquima y del colénquima que dan lugar a células que se distribuyen de manera desigual hacia dentro o hacia fuera del individuo. Las capas más externas son más numerosas, sus células se suberifican y mueren formando el súber o corcho. Hacia dentro las células están vivas en una disposición apilada formando la felodermis.
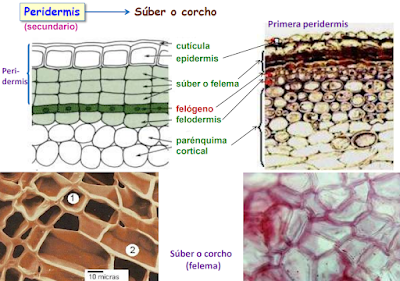
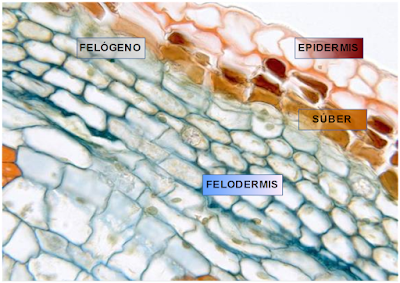
El súber o corcho está formado por varias capas de células muertas y se sitúa directamente por debajo de la epidermis. Se desarrolla a partir del felógeno que, por división de las células de su única capa, origina hacia la periferia células aplanadas que sufren la suberificación de las paredes por depósito de una sustancia llamada suberina que conduce a la degeneración del protoplasma y a la muerte posterior. La suberina es impermeable a los líquidos y a las sustancias gaseosas, por lo que el súber interfiere de alguna manera en la transpiración y el intercambio de gases. Por otra parte, el súber es un moderador térmico, lo que constituye la principal defensa del vegetal contra los cambios bruscos de temperatura. Por último, debido a la resistencia de este tejido frente a las enzimas liberadas por distintos parásitos, el súber se presenta como una barrera hacia los mismos.
El súber puede ser retirado y la planta sobrevive (salvo que se vea atacada por patógenos como los hongos) formando nuevo súber posteriormente.

Como el súber impide el intercambio gaseoso presenta las lenticelas que son aberturas en el súber donde se disponen células parenquimáticas vivas de paredes finas y sin suberificar que dejan huecos entre ellas para facilitar el intercambio de gases y la transpiración.
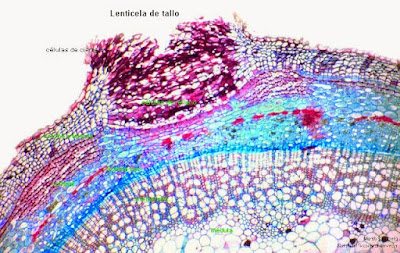


La felodermis es una capa concéntrica formada por la división del felógeno hacia el interior. Sus células presentan un aspecto irregular y a diferencia del súber, tienen un protoplasma vivo. El número de células de esta capa es muy inferior a las del súber y pueden estar provistas de cloroplastos.
2.2.4. Tejidos vegetales conductores (xilema y floema)
Estos tejidos se encargan de distribuir los nutrientes por todas las partes del vegetal. Se sitúan en el centro de la raíz y del tallo, formando estructuras lineales que los recorren en toda su longitud. También se introducen en las hojas formando los nervios.
Además de su función primordial en el reparto de los nutrientes por la planta son muy importantes como elementos de sostén y para la comunicación entre las distintas partes de la planta (gracias a las hormonas vegetales).
Los tejidos conductores son tubos que se asocian formando haces o grupos. Además de las células principales del tejido, llevan también otras células acompañantes que realizan distintas funciones. En las plantas se encuentran dos clases de tejidos conductores:
a) El xilema es un tejido complejo formado por varios tipos celulares. Su función es la conducción de agua y minerales desde la raíz hasta las hojas.
b) EI floema, al igual que el xilema, es un tejido complejo que se extiende por todo el cuerpo de la planta, como un sistema continuo, desde las hojas hasta el ápice de la raíz y gracias a este tejido, las sustancias elaboradas y otras sustancias disueltas incluyendo las sales minerales, son distribuidas a todas las partes de la planta.







2.2.4.1. El xilema
El xilema, también llamado leño, se encarga del transporte y reparto de agua y sales minerales provenientes fundamentalmente de la raíz al resto de la planta, aunque también transporta otros nutrientes y hormonas vegetales. Es también el principal elemento de soporte mecánico de las plantas, sobre todo en aquellas con crecimiento secundario (la madera es básicamente xilema).
El xilema que se diferencia en el cuerpo primario de la planta se denomina xilema primario, y su precursor es el procambium. Este xilema es el definitivo en aquellas plantas sin un crecimiento secundario.
Consta de una parte que se diferencia temprano en las partes primarias de la planta que aún no han completado su desarrollo y diferenciación, es el protoxilema(posteriormente sustituido); y otra, el metaxilema que se inicia generalmente en el cuerpo primario, de la plantas que todavía se halla en crecimiento, pero por lo general madura cuando se ha completado el alargamiento de las plantas.

El xilema formado después de terminado el crecimiento primario de la planta se llama xilema secundario, y es formado por el cambium vascular.
La estructura característica del xilema secundario es la existencia de dos sistemas de elementos, que difieren en la orientación de sus células: uno es horizontal y el otro es vertical.
Sistema vertical o longitudinal o axial: son células o filas de células con el eje mayor orientado longitudinalmente, formado por elementos conductores no vivos y células parenquimáticas vivas.
Sistema horizontal o transversal o radial: son hileras de células orientadas radialmente, formado por células vivas principalmente, las células parenquimáticas de los radios medulares.
Las células vivas de los radios y del sistema axial se encuentran generalmente en conexión formando un sistema continuo.
El xilema secundario producido durante un período de crecimiento constituye una capa , que en corte transversal de tallo se llama anillo de crecimiento. Si se observa a simple vista tiene una parte clara, que es el leño temprano o de primavera, menos denso, con células de mayor diámetro y una parte oscura, que es el leño tardío, sus células son pequeñas y de paredes más gruesas. Esto ocurre generalmente en especies que viven en regiones templadas.

En el xilema nos encontramos cuatro tipos celulares principales: Los elementos de los vasos o tráqueas y las traqueidas constituyen los elementos conductores o traqueales; las células parenquimáticas, que funcionan como células de almacenamiento o comunicación, y las células de sostén, que son las fibras de esclerénquima y las esclereidas.
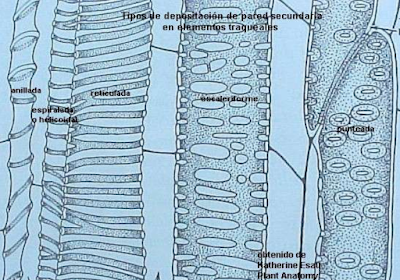
A) Elementos de los vasos o tráqueas; son células con una pared secundaria gruesa, dura y lignificada. En su madurez mueren y su citoplasma se elimina.
Son células de mayor diámetro y más achatadas que las traqueidas.

Se unen longitudinalmente unas a otras para formar tubos llamados vasos. Por ellos el agua circula vía simplasto (por el interior de las células), y pasa de una célula a la siguiente por las perforaciones que se encuentran en sus paredes transversales (situadas en ambos extremos de la célula), denominadas placas perforadas.
Además, el agua y sustancias disueltas pueden atravesar las punteaduras areoladas situadas en las paredes laterales de la célula y pasar a otras células del xilema. Los elementos de los vasos son el principal tipo celular conductor del xilema en las angiospermas.

B) Traqueidas; son el otro elemento conductor que aparece en las plantas vasculares. Las pteridofitas y gimnospermas sólo poseen este tipo traqueal como célula conductora, mientras que las angiospermas poseen tanto traqueidas como elementos de los vasos.
Las traqueidas son células alargadas, estrechas y fusiformes. El agua circula por ellas y pasa de unas a otras vía simplasto atravesando las punteaduras areoladas. Estas punteaduras se encuentran sobre todo en las paredes de ambos extremos de la célula, las cuales se solapan entre células contiguas.
En general su capacidad para conducir agua es menor que la de los elementos de vasos ya que no poseen placas perforadas. Además, tienen paredes celulares más gruesas y un menor volumen interno para la conducción que los elementos de los vasos.





2.2.4.2. El floema
El floema, llamado también líber o tejido criboso, es un tejido de conducción formado por células vivas. Su principal misión es transportar y repartir por todo el cuerpo de la planta las sustancias carbonadas producidas durante la fotosíntesis, o aquellas sustancias movilizadas desde los lugares de almacenamiento, además de otras moléculas como las hormonas vegetales.
El floema está formado por dos tipos de células conductoras: los tubos o elementos cribosos y las células cribosas. Ambos tipos celulares son células vivas, aunque sin núcleo, y tienen la pared primaria engrosada con depósitos de calosa.


A) tubos o elementos cribosos; son típicos de las angiospermas. Son células individuales achatadas que se disponen en filas longitudinales y que se comunican entre sí mediante placas cribosas localizadas en sus paredes transversales o terminales.
Las placas cribosas contienen poros de gran tamaño que comunican los citoplasmas de las células vecinas. Además, poseen áreas cribosas en las paredes laterales que son depresiones en la pared primaria con poros que atraviesan la pared completamente. Éstas sirven para comunicarse con otros tubos cribosos contiguos y con las células parenquimáticas especializadas que los acompañan llamadas células acompañantes o anexas.




B) células cribosas; son típicas de gimnospermas y pteridófitas. Son células largas y de extremos puntiagudos, comunicándose entre sí lateralmente mediante grupos de campos de poros primarios que forman las áreas cribosas. Sin embargo, no poseen placas cribosas. Se relacionan funcional y morfológicamente con una célula parenquimática especializada llamada célula albuminosa.

Dentro de los elementos no conductores se encuentran las células parenquimáticas, siendo las más abundantes las denominadas células acompañantes (encargados de nutrir a los elementos conductores que no tienen núcleo y poco citoplasma en las angiospermas) y las células albuminosas (que hacen lo mismo en las gimnospermas). Como células no conductoras también se pueden encontrar células de soporte asociadas al floema, entre las que se encuentran las fibras de esclerénquima y las esclereidas.
Las fibras de esclerénquima y las esclereidas se encuentran asociadas al floema con una función de protección y soporte.

El floema primario es el primer tipo de floema que aparece en los órganos en desarrollo, aparece primero como protofloema y más tarde como metafloema.
El protofloema es el primer floema que aparece y se forma a partir del meristemo procámbium. El protofloema contiene elementos cribosos poco desarrollados en las angiospermas, mientras que en las gimnospermas y pteridófitas posee células cribosas, también poco desarrolladas. Las células acompañantes son muy raras o ausentes.
El metafloema sustituye rápidamente al protofloema, normalmente cuando termina la elongación del órgano, y se origina también a partir del procámbium. Contiene tubos cribosos y células cribosas de grosor y longitud mayores que en el protofloema, y siempre tienen células acompañantes. Aquí aparecen las placas cribosas en los tubos cribosos. Este tejido es funcional en las plantas con crecimiento primario.
El floema secundario se forma a partir del meristemo cámbium vascular en las plantas con crecimiento secundario. En este tipo de floema los elementos conductores están muy desarrollados, así como las células acompañantes, y aparecen tanto el parénquima axial como el radiomedular. Las células del floema secundario, al contrario que en el xilema, no depositan pared celular secundaria, y son células vivas. Sin embargo, el citoplasma de los elementos cribosos puede carecer de núcleo, microtúbulos y ribosomas, y el límite entre la vacuola y el resto del citosol no es claro. En los árboles en crecimiento hay muy poco floema secundario activo implicado en la conducción de nutrientes.








Son estructuras situadas en la epidermis o en el súber. Contiene glándulas de distinta estructura que forman sustancias que secretan al exterior. Estas sustancias pueden ser productos de desecho o bien son sustancias útiles para la planta. Las sustancias principales que expulsan las plantas son:
a) Aromas y Aceites.
b) Venenos (extramonio).
c) Sustancias urticantes.
d) Látex y resinas.


Los tejidos secretores se encuentran diseminados en los más distintos tejidos, tanto primarios como secundarios, y estos tejidos de acuerdo a la distribución que tienen en la planta se clasifican en:
A) Tejidos secretores epidérmicos: Estos tejidos son células, o conjunto de células, que se encuentran en la epidermis de flores, tallos y hojas de muchas plantas. Las células secretoras activas, tienen protoplastos densos y elaboran sustancias de la familia de los terpenos, como: Aceites esenciales, bálsamos, resinas, sustancias que pasan del citoplasma a través de la pared celular para acumularse entre la pared celulósica y la cutícula; su separación definitiva tiene lugar por ruptura de la cutícula.

B) Tejidos secretores internos: Son aquellos tejidos que se encuentran en el interior de otros tejidos; pueden estar constituidos por células aisladas simples, se encuentran diseminadas en el tejido medular o también pueden agruparse alrededor del espacio intercelular y formar un canal o bolsa, en donde vierten su secreción al interior.


C) Tejidos laticíferos: Son tejidos secretores internos, constituidos por células o serie de células que se encuentran unidas formando sistemas complejos de forma tubular, en los que es muy difícil reconocer los límites de las células individuales. Reciben este nombre porque estos tejidos contienen un liquido lechoso, llamado látex. Generalmente los laticíferos se hallan distribuidos por toda la planta, pero también están limitados a ciertos tejidos (mesófilo de las hojas), partes aéreas de la planta, en raíces, incluso estan asociados al floema.

D) Conductos resiníferos: Son típicos de las coníferas y se pueden encontrar entre las células del parénquima cortical tanto en las hojas y como en los tallos, o dispuestos entre las células del xilema secundario. Son estructuras alargadas que delimitan una cavidad central revestida por un epitelio secretor. Las células son en general de paredes finas y secretan a la cavidad central una mezcla de sustancias en las que abundan los terpenoides y que constituye la resina. Se cree que la resina protege a la planta del ataque de hongos y de insectos.

3. Organografía vegetal
Las plantas tienen tres órganos vegetativos que conforman su cuerpo: la raíz, el tallo y las hojas. También presenta órganos reproductores como la flor o el fruto.




3.1. La raíz
La raíz es el órgano de la planta que se localiza frecuentemente bajo tierra, ya que presenta geotropismo positivo.
La raíz es la primera estructura embrionaria que se desarrolla a partir de la germinación de la semilla. La radícula es una estructura inicialmente poco diferenciada que dará origen a la raíz primaria cubierta por la caliptra, que actúa como protector apical.
El eje principal de las plantas está constituido por el tallo y la raíz. La unión de ambas estructuras no presenta una diferenciación evidente, ya que los tejidos vasculares están incluidos en el tejido fundamental.
La morfología de la raíz es más sencilla debido a su hábitat bajo la tierra. En las raíces no hay presencia de nudos, yemas, estomas, ni producción de clorofila, con algunas excepciones de raíces adaptadas a condiciones especiales.
Esta estructura esta encargada de la absorción y transporte de agua y nutrientes almacenados en el suelo. Los pelos absorbentes captan estos elementos -savia bruta-, que son transportados hacia el área foliar donde son transformados en el proceso de fotosíntesis.
Así mismo, las raíces sujetan las plantas al terreno impidiendo su desprendimiento por medio de agentes externos. En otros casos las raíces actúan como estructuras de almacenamiento o reserva de elementos nutricionales, por ejemplo, la remolacha.
3.1.1. Partes de la raíz
a) Cofia o caliptra; es la envoltura de recubrimiento exterior que protege la punta de la raíz y contribuye con la penetración hacia el interior del suelo. Se origina a partir del dermatógeno y meristemo derivado protodermis -dicotiledóneas-, o en el caliptrógeno -monocotiledóneas-.
La caliptra está constituida por células con abundante contenido de almidón y dictiosomas, además de mucílagos que favorecen el avance de la raíz en el suelo. Su función es básicamente la protección de la zona meristemática.
b) Zona meristemática; está conformada por tejidos meristemáticos que tienen la capacidad de dividirse por mitosis y dar origen al meristemo derivado: meristemo fundamental, protodermis y procámbium.
c) Zona de crecimiento; en la zona de crecimiento o alargamiento ocurre el alargamiento de las células de los meristemos derivados, es el lugar donde ocurren las divisiones celulares y se inicia el proceso de diferenciación de los tejidos.
d) Zona de diferenciación o pilífera; en esta zona es común observar gran cantidad de finos pelos radiculares cuya función es la absorción de los nutrientes, agua y sales minerales. Además, a partir de este punto se observan los tejidos primordiales que constituyen la estructura primaria de la raíz.
e) Zona de ramificación; comprende la zona desde el cuello hasta la zona pilífera. Es la zona donde se desarrollan y crecen las raíces secundarias o laterales. Finaliza en el lugar donde se origina el tallo o cuello de la planta.
f) Cuello; es el lugar donde la raíz se une con el tallo.

3.1.2. Estructura interna de la raíz
La estructura interna básica de la raíz consiste en los siguientes elementos (desde el exterior hacia el interior):
a) una capa externa, la epidermis, con pelos radicales absorbentes.
b) el córtex, formado por tejido parenquimático, sin clorofila, que puede acumular almidón.
c) la endodermis, llamada banda de Caspary, rica en suberina y con función impermeable, que hace que el agua fluya desde el exterior al interior de la raíz y no al contrario.
d) el cilindro vascular, constituido por el xilema y el floema.

Podemos distinguir entre dos tipos de estructura:
a) Estructura 1ª de la raíz; se produce el crecimiento en longitud de la raíz a partir de los meristemos apicales y la sucesiva diferenciación del resto de los tejidos.





b) Estructura 2ª de la raíz; se produce el crecimiento en grosor de la raíz en las plantas plurianuales.





3.1.3. Tipos de raices



El tallo es la parte aérea de la planta en la que se desarrollan las hojas, aunque en algunas especies puede ser subterráneo. Su función, además de proporcionar el soporte a la planta, es transportar la savia bruta desde la raíz hasta los órganos fotosintéticos, y la savia elaborada mediante la fotosíntesis a todo el vegetal.


Podemos distinguir entre dos tipos de estructura:
a) Estructura 1ª del tallo; se produce el crecimiento en longitud del tallo. Es algo más complejo que en las raíces porque tiene que dar lugar a los primordios foliares y a las ramas en los nudos.






b) Estructura 2ª del tallo; se produce el crecimiento en grosor del tallo. Es característico de las plantas dicotiledóneas plurianuales y de las gimnospermas.









3.2.3. Tipos de tallos
Las hojas son órganos generalmente aplanados derivados de un meristemo caulinar apical. Son los órganos fotosintéticos por excelencia de las plantas gracias a la enorme cantidad de cloroplastos que poseen sus células. Además, las hojas son las principales responsables de controlar la transpiración para evitar la pérdida excesiva de agua. El diseño y la distribución de las hojas en el cuerpo de la planta se pueden explicar si tenemos en cuenta estas funciones. Durante la evolución, las plantas vasculares inventaron las hojas probablemente a partir de ramas.
3.3.1. Morfología de las hojas
Las hojas se pueden dividir anatómicamente en dos partes: limbo y peciolo.

Las hojas se pueden dividir anatómicamente en dos partes: limbo y peciolo.
El limbo es la parte de la hoja encargada de realizar la fotosíntesis y regular la transpiración. Aquí se encuentran la mayoría de los estomas y del parénquima clorofílico de la planta.
El limbo posee dos superficies, una superior, denominada haz. y otra inferior, denominada envés. La superficie que normalmente queda expuesta al Sol es el haz, mientras que el envés es la superficie que queda oculta. Se denomina contorno al borde del limbo y puede ser muy variado en su forma.
El peciolo es una estructura más o menos larga y cilíndrica que une el limbo al tallo a nivel de los nudos. En el ángulo agudo que se forma en el punto de unión entre el tallo y el peciolo se localizan las yemas axilares de las que partirán nuevas ramas. Hay hojas sésiles, que carecen de peciolo, donde el limbo se une directamente al tallo.

El tamaño de las hojas es variable y en general una hoja pequeña se asocia a lugares con una mayor altitud, poca lluvia, pocos nutrientes en el suelo, y a lugares calientes y secos. La exposición a la luz solar es otro factor importante que afecta al tamaño y grosor de las hojas, incluso en una misma planta. Las hojas con más exposición solar son más pequeñas y más gruesas, sobre todo por el desarrollo del parénquima, pero también tienen un sistema vascular y una epidermis más desarrollados que las hojas denominadas de sombra.
En función de la complejidad del limbo se puede dividir a las hojas en simples y compuestas. Las simples tienen un limbo continuo y sin dividir, mientras que las compuestas poseen varias subunidades, denominadas foliolos, cada una de ellas asemejándose a una hoja distinta. Distinguir una hoja de un foliolo es posible por la presencia o no de yemas axilares, sólo las hojas las tienen.

La variabilidad de la forma del limbo también es enorme y viene genéticamente condicionada por la especie, lo cual es el resultado de una adaptación evolutiva a su medio ambiente, incluyendo a los herbívoros.


También distinguimos varios tipos de hojas según la forma del borde del limbo:

Los haces vasculares de la hoja se denominan también nervios o venas, y su manera de organizarse se denominada nerviación o venación de la hoja.
En las dicotiledóneas suele haber un nervio principal que se va ramificando y en cada ramificación disminuye el diámetro del nervio. A este patrón de ramificación se le denomina reticular. El nervio principal recorre el eje central de la hoja, desde el peciolo hasta la punta de la hoja. A este nervio más el tejido que le rodea se les denomina costilla.
En las monocotiledóneas los nervios corren paralelos al eje principal de la hoja y son del mismo diámetro. A este tipo de nerviación se le llama paralela. Sin embargo, existen vasos conductores de pequeño calibre que conectan a los grandes y paralelos.


En la base de las hojas se desarrollan unas estructuras a modo de pequeñas hojas o escamas denominadas estípulas, pero en las monocotiledóneas la escama se dilata para formar vainas que abrazan al tallo.
Algunas hojas modifican enormemente su desarrollo y forman estructuras que no están estrictamente relacionadas con la fotosíntesis. Así, algunas hojas se asocian a las flores para formar las brácteas que rodean a los pétalos, pueden formar espinas como en algunos espinos (no confundir con las espinas de las zarzas que son derivadas del tallo), sirven para atrapar insectos en las plantas carnívoras, etcétera.

La disposición de las hojas en el tallo principal y en las ramas laterales se da en las plantas que tienen una vida corta de una temporada. Pero en las plantas que viven varios años las hojas crecen en la parte de las ramas laterales que han crecido durante ese año o hace pocos años. Las hojas caen en los árboles caducos, pero en los perennes pueden durar varios años. Independientemente de eso las hojas nuevas crecen en los brotes nuevos. La organización de las hojas puede ser en espiral cuando las hojas aparecen alternando la posición y la altura a modo de hélice, opuestas cuando hay dos hojas al mimo nivel y una a cada lado del tallo, o verticiladas cuando nos encontramos tres o más hojas a una misma altura en el tallo. Se llama filotaxia a la organización de las hojas.

3.3.2. Estructura interna de las hojas

En la superficie del haz hay una epidermis cutinizada con capas muy gruesas de cutina y ceras, y, por lo general sin estomas, mientras que en la superficie del envés hay una epidermis más delgada y con gran densidad de estomas.
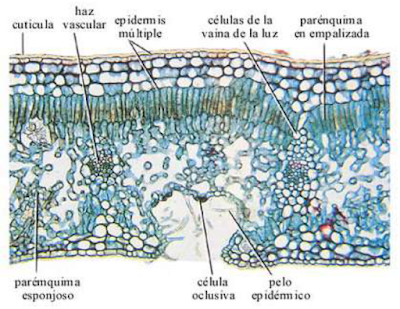
En la epidermis de las hojas de muchas plantas hay algunas células que se diferencian para transformase en pelos o tricomas. Los tricomas pueden ser unicelulares o multicelulares, y pueden tener diversas funciones como protección, glandulares, evitar la pérdida de agua, etcétera. Las hojas que carecen de tricomas se denominan glabras y las que los tienes pubescentes. En muchas monocotiledóneas hay unas células diferenciadas en el epidermis denominadas buliformes cuya función es desenrrollar y extender la hoja cuando hay humedad.

Entre las dos capas epidérmicas nos encontramos con el mesófilo, que es tejido parenquimático. Suelen distinguirse dos tipos presentes en la misma hoja: parénquima en empalizada (o clorofílico) y el parénquima lagunar (o esponjoso, que se suele también llamar aerénquima).
Los espacios intercelulares están conectados con los estomas y son importantes para el trasiego de gases necesarios para la fotosíntesis.
En muchas hojas hay esclerénquima y colénquima localizados en el margen de la hoja, bajo la epidermis, formando una capa denominada hipodermis. La hipodermis, además de sostén, contribuye a evitar la pérdida de agua. También hay esclerénquima rodeando a los vasos conductores, proporcionan sostén a los tejidos de la hoja. Las hojas de las plantas acuáticas tienen muy poco esclerénquima y colénquima o carecen de ellos. El mesófilo puede contener conductos secretores como los resiníferos de los pinos, cavidades de aceites como en el eucalipto, laticíferos, etcétera.
Las hojas poseen vasos conductores, denominados nervios, para llevar agua y sales a las células fotosintetizadoras, y recoger los productos orgánicos y repartirlos por el resto de la planta. Los vasos están formados por xilema y floema, además de tejido parenquimático alrededor. El xilema normalmente está orientado hacia el haz, mientras que el floema lo está hacia el envés.

La flor es el órgano reproductivo de las espermatofitas (plantas con semilla) que está representado por gimnospermas y angiospermas. Ambos grupos de plantas tienen flores pero en el caso de las gimnospermas las flores son en realidad inflorescencias que no van a dar lugar a un fruto. Por el contrario las angiospermas presentan flores típicas que tras la fecundación formarán semillas encerradas en frutos. Vamos a estudiar la estructura de la flor de angiospermas por representar al grupo de plantas más abundantes y diversas, y las más fáciles de observar.
Las flores aparecen durante la época reproductiva de las plantas mediante un proceso denominado floración. Como todos los órganos de la planta, las flores se originan a partir de la actividad meristemática. En realidad una flor podría considerarse como una porción de tallo altamente modificada donde las hojas sufren cambios drásticos para convertirse en las diferentes partes de la flor.
La parte que une la flor al tallo se denomina pedúnculo, y las flores que carecen de él se denominan sésiles. Cuando el pedúnculo sostiene a un grupo de flores, como si fuera un ramo, a la estructura que une a cada flor con el pedúnculo se le denomina pedicelo. El pedúnculo o pedicelo, forman una estructura terminal denominada receptáculo, desde donde parten las otras estructuras de la flor.

Una flor típica consta de dos partes:
a) Perianto; es la parte estéril de la flor. Tiene una función protectora o favorecedora de la fecundación. Está compuesta por el cáliz (conjunto de sépalos) y por la corola (conjunto de pétalos). Los sépalos suelen ser de menor tamaño, verdes (tienen muchos cloroplastos) y su función es sostener y proteger a la flor. Los pétalos suelen ser de mayor tamaño, coloreados y su función es atraer a los polinizadores.
b) Parte reproductora: está formada por el androceo (los estambres), que constituye la parte masculina de la flor, y por el gineceo (el pistilo/carpelos), que es la parte femenina.
Un estambre típico consta de un filamento en cuyo extremo distal se encuentra la antera. Ésta última es un cuerpo alargado con una invaginación medial que la divide en dos lóbulos o tecas, conteniendo cada una de ellas dos sacos polínicos. En estos sacos ocurre la microesporogénesis o formación de microsporas que se convierten en el gametofito masculino o grano de polen.
El gineceo está formado por uno o por un conjunto de pistilos, cada uno de los cuales está formado por una o varias hojas modificadas denominadas carpelos, que se doblan sobre sí mismas formando esa estructura típica en botella.
La base dilatada del pistilo es el ovario, que contiene a los rudimentos seminales. En los rudimentos seminales se encuentran los óvulos, se producirá la fecundación y a partir de ellos se desarrollarán las semillas. Dentro de los rudimentos seminales se encuentran los gametofitos femeninos.
El tubo del pistilo se denomina estilo. La parte superior y pegajosa del pistilo se denomina estigma.

El receptáculo es donde se insertan los estambres, pétalos y sépalos de la flor.
La semilla se forma a partir del rudimento seminal, localizado en el ovario de las flores, tras producirse la fecundación por los granos de polen.
Se pueden distinguir diferentes partes en una semilla:
a) Embrión; está compuesto por un eje embrionario (en cuyos extremos se encuentran una radícula y una plúmula, y por uno o dos cotiledones.
Los cotiledones pueden almacenar sustancias de reserva para la germinación y entonces suelen tener un aspecto carnoso. La porción del eje que queda apical al punto de inserción de los cotiledones, hacia la plúmula, se denomina epicótilo, mientras que la que queda por debajo se denomina hipocótilo.
b) Endospermo secundario; es un tejido nutritivo que se encuentra a un lado del embrión o rodeándolo. El endospermo es un tejido de reserva que proporciona nutrientes al embrión y a las primeras fases del desarrollo de la planta. Las células nutricias almacenan granos de almidón o de proteínas.
c) Cubiertas protectoras; la formación de la cubierta está inhibida antes de la fecundación y la fecundación elimina esta inhibición permitiendo el desarrollo de la cubierta. La cubierta en su conjunto se denomina epispermo o cubierta seminal.
Una característica de la cubierta es que debe mantener protegida a la semilla del exterior, pero al mismo tiempo debe sentir el ambiente para permitir la germinación cuando las condiciones son adecuadas.


Después de la fecundación, a la vez que el rudimento seminal se convierte en semilla, comienza la transformación de las paredes del ovario para formar el fruto. Se puede considerar al fruto como el órgano que contiene a la semilla hasta el final de su maduración. Es un ovario maduro con algunas partes añadidas como el receptáculo, cáliz y brácteas. El fruto no sólo protege sino que también ayuda a la dispersión de la semilla, bien de forma activa o pasiva. Algunas semillas no pueden germinar a no ser que hayan pasado antes por el tracto digestivo de un animal.
Podemos distinguir:
a) Un fruto simple está compuesto por un carpelo, o por varios carpelos fusionados de una misma flor. Por ejemplo, el melocotón o el tomate.

b) Un fruto múltiple o compuesto es aquel que se forma de la fusión de varias flores, tales como el higo.

c) Los frutos pueden ser dehiscentes, si se abren en algún momento dejando libres a las semillas, o indehiscentes si las semillas no quedan libres hasta que el fruto se descompone totalmente.
d) Los frutos secos son aquellos cuyo pericarpo es membranoso o coriáceo, poco desarrollado y poco hidratado.


e) Los frutos carnosos son aquellos cuyo pericarpo tiene una consistencia carnosa, es suculento y está muy hidratado.

Este comentario ha sido eliminado por el autor.
ResponderEliminar