En 1665, el naturalista inglés Robert Hooke observó una fina lámina de corcho con un microscopio muy básico y distinguió en ella unas celdillas o huecos a las que denominó células. Sin embargo, realmente estaba observando las paredes celulares de células muertas, cuyo interior estaba vacío.


Poco después, en 1674, el holandés Antón van Leeuwenhoek construyó un microscopio rudimentario, de una sola lente, con el que observó gotas de agua, sangre, esperma, etc. Describió con gran detalle los seres y las células que observó, a los que denominó «animáculos».


Podéis leer unos artículos muy interesantes al respecto aquí
La historia de van Leeuwenhoek
Los microscopios de van Leeuwenhoek
Posteriores avances en el desarrollo del microscópico y diversas investigaciones llevaron a la conclusión de que las células no estaban vacías y que en su interior se podían observar estructuras. Por lo tanto los avances en microscopía han sido determinantes para poder avanzar en el estudio de las células.
1. La teoría celular
La teoría celular se debe a dos científicos alemanes, el botánico Mathias Schleiden y el zoólogo Theodor Schwann. En 1838, Schleiden señaló por primera vez que las plantas se componen de células. Al año siguiente, Schwann extendió esta generalización a los animales. La teoría celular no tardó en imponerse, pues agrupó un conjunto de datos que ya gozaban de consenso en la comunidad científica y desde entonces se acepta que la célula es la unidad básica de todos los organismos vivos.

En el año 1855, Rudolf Virchow amplió la teoría celular y afirmó que las células solo surgen por división de otras células preexistentes, contradiciendo así la teoría (que aún entonces tenía muchos adeptos), de que las células pueden surgir por generación espontánea de la materia inanimada.

Durante el siglo XX, la teoría celular fue reafirmada y ampliada y es hoy uno de los conceptos unificadores más importantes de la biología. En su formulación actual, la teoría celular enuncia:
a) Los seres vivos están formados por células y productos celulares (unidad estructural).
b) Las células se originan a partir de otras células (unidad reproductiva).
c) Las reacciones químicas del organismo vivo tienen lugar dentro de células (unidad fisiológica).
d) Las células contienen la información hereditaria de los organismos que integran y esta información se transmite de la célula madre a la célula hija (unidad genética).
Todas aquellas características que se hacen evidentes en un organismo complejo y nos permiten reconocerlo como un ser vivo, están presentes en cada una de las células que lo componen.
2. Tipos de células
La célula es la estructura viva más sencilla que se conoce, es decir que es capaz de realizar las tres funciones vitales, que son nutrirse, relacionarse y reproducirse. Todas las células conocidas constan de:
a) Una membrana plasmática que las individualiza del medio externo y que sirve de superficie de intercambio entre el interior y el exterior celular.
b) Un citoplasma en él que se puede diferenciar un medio líquido denominado citosol o hialoplasma y una serie de estructuras denominadas orgánulos celulares.
c) Un material genético en forma de ADN que contienen la información genética, es decir la información de como es y como funciona la célula. Según que las moléculas de ADN estén dispersas en el citosol o rodeadas de una membrana especial formando una estructura denominada núcleo, se diferencian dos tipos principales de células: las procariotas y las eucariotas.
2.1 La célula procariota (Dominios Bacteria y Archaea)
Son las células que no tienen núcleo, es decir son las que presentan su ADN más o menos condensado en una región del citoplasma pero sin estar rodeado de una membrana. El ejemplo más importante de células procariotas son las bacterias.
Son células mucho más sencillas y pequeñas que las eucariotas (0,5 a 5 μm).
Suelen presentar una gruesa pared celular con dos tipos básicos que permite establecer dos grandes grupos de bacterias (gram+ y gram-). En ocasiones está pared está recubierta por una cápsula o glucocálix que les permite resistir la desecación y el ataque de elementos como los anticuerpos (las bacterias con cápsula son más virulentas).
Pueden presentar flagelos (normalmente 1 o 2) para el desplazamiento, fimbrias para adherirse a determinadas superficies y pilis (o pelos) para intercambiar información genética con otras bacterias (fenómenos parasexuales). Carecen de citoesqueleto y de movilidad intracelular.
Como orgánulos encontramos ribosomas 70S (más pequeños que los eucariotas) y mesosomas, expansiones de la membrana celular para aumentar la superficie (aunque muchos autores los consideran artefactos causados por las técnicas usadas para su observación microscópica).
El ADN se encuentra formando un cromosoma circular que no está limitado por ninguna membrana: se le denomina nucleoide porque suele ocupar una posición central. También encontramos pequeñas moléculas de ADN circular llamadas plásmidos.





2.2 La célula eucariota (Dominio Eukarya)
La célula eucariota está constituida por tres partes denominadas membrana plasmática, citoplasma (con numerosos orgánulos especializados) y núcleo (donde se encuentra albergado el ADN). Está mayor compartimentalización es posible gracias a un conjunto de membranas biológicas situadas en el citoplasma e independientes de la membrana plasmática. Esto permite a las células eucariotas una mayor eficiencia y, por lo tanto, alcanzar tamaños mucho mayores (10-100 μm).
Estas células son las que han originado a todos los organismos pluricelulares al formar los diferentes tejidos.
Se diferencian dos tipos principales: las células eucariotas animales o heterótrofas y las células eucariotas vegetales o autótrofas.
a) Células animales o heterótrofas; se caracterizan por no presentar pared celular o, en el caso de los hongos, que no sea nunca de celulosa, tener vacuolas muy pequeñas, carecer de cloroplastos y presentar centrosoma (orgánulo relacionado con la organización de cilios y flagelos). Su forma suele ser globular aunque puede cambiar en función del tejido.


b) Células vegetales o autótrofas; se caracterizan por presentar una pared gruesa de celulosa situada en el exterior de la membrana plasmática, por tener grandes vacuolas y cloroplastos y por no tener centrosoma. Su forma suele ser prismática.


Las principales diferencias entre las células animales y vegetales son:
a) En general, las células vegetales suelen tener forma prismática, mientras que las animales presentan formas muy diversas (estrellada, alargada, globular, aplanada, etc).
b) Las células vegetales están recubiertas de una pared celular rígida formada por celulosa que contribuye a mantener la forma y el volumen de la célula.
c) En las vegetales existen los cloroplastos, orgánulos que acumulan clorofila, pero también existen otros tipos de plastos que acumulan otras sustancias, por ejemplo los leucoplastos que acumulan almidón (son muy abundantes en las patatas) y los cromoplastos que acumulan sustancias coloreadas (se encuentran en los pétalos de las flores).
d) En las células animales tenemos centriolos, pero no en las vegetales.
e) En las células vegetales hay un gran vacuola central, llamada vacuola de turgencia, que ocupa la mayor parte de la célula permitiendo que se mantenga su forma. Sin embargo, en las animales suelen ser más pequeñas y su misión es la reserva de nutrientes
f) El reparto del citoplasma en la división celular se realiza en las células animales por estrangulamiento, y en las vegetales por el crecimiento de un tabique de separación denominado fragmoplasto.
3. La teoría endosimbionte
Esta teoría sobre el origen de la célula eucariota se contrasta con la teoría antes predominante (teoría autógena) que defendía que la célula eucariota se formaba por el aumento de tamaño y complejidad de algunas células procariotas que conllevaba el desarrollo del sistema endomembranoso y, posteriormente, de los orgánulos membranosos.
La teoría endosimbiótica seriada que planteó Lynn Margulis afirmaba que la célula eucariota se había formado por la simbiosis permanente entre diferentes tipos de procariotas. Según Margulis este proceso habría ocurrido en tres fases:
a) En una primera incorporación se habría incorporado a un procariota de gran tamaño una bacteria nadadora (espiroqueta) que formaría el flagelo y una primera célula con una membrana interna. Este sería el primer eucariota ancestral.
b) En una segunda incorporación se habría incorporado una bacteria respiradora de oxígeno a un eucariota anaerobio. Esto terminaría dando lugar a los eucariotas animales o los hongos y a las mitocondrias.
c) En una tercera incorporación se habría incorporado una bacteria fotosintética a una primitiva eucariota animal originando las primitivas células vegetales.

Hoy en día se acepta de forma prácticamente total el origen endosimbiótico de mitocondrias y cloroplastos dado que numerosas pruebas apuntan a ello. Sin embargo el origen del flagelo o de las membranas internas es mucho más discutido dado que su estructura no concuerda con las observadas en los eucariotas.
Argumentos a favor de la teoría endosimbiótica
a) El tamaño de las mitocondrias es similar al tamaño de algunas bacterias.
b) Las mitocondrias y los cloroplastos contienen ADN bicatenario circular similar al de procariotas.
c) Las mitocondrias y los cloroplastos están rodeados por una doble membrana y la interna es muy similar a la bacteriana.
d) Las mitocondrias y los cloroplastos se dividen por fusión binaria al igual que los procariotas.
e) Las mitocondrias y los cloroplastos tienen material genético propio e independiente y sintetizan la mayor parte de las proteínas que usan estos orgánulos.
f) En mitocondrias y cloroplastos encontramos ribosomas 70S (similares a los procariotas) mientras que en el citoplasma encontramos ribosomas 80S.
4. Las envolturas celulares
Las envolturas celulares son capas que separan el medio interno del exterior. Son: la membrana plasmática, que poseen todas las células (eucariotas y procariotas) y las membranas de secreción (que pueden faltar). Son membranas de secreción: la matriz extracelular (de animales), la pared vegetal (de vegetales) y la pared bacteriana (en bacterias).
4.1 La membrana plasmática
La membrana plasmática constituye el límite entre el citoplasma y el medio en el que se encuentra la célula y entre los orgánulos celulares y el citosol (hialoplasma), de manera que las biomembranas dividen al interior de la célula en numerosos compartimentos. De esta manera se explica que las membranas puedan constituir el 60% del citoplasma celular.
Posee un espesor de 75 Ǻ. Al microscopio electrónico se presenta como una triple capa. Dos bandas oscuras externas de 20 Ǻ separadas por una interna de color claro de 35 Ǻ. En ocasiones, las láminas externas más oscuras pueden tener espesores diferentes.

Cualquier tipo de célula presenta este tipo de membrana de tres capas. Debido a su carácter generalizado se le denomina unidad de membrana (o membrana unidad). También se encuentra formando la envoltura exterior de muchos orgánulos como los cloroplastos, mitocondrias o vacuolas.
4.1.1. Composición química de la membrana plasmática
Las membranas biológicas son conjuntos laminares constituidos aproximadamente por un 40 % de lípidos y un 60 % de proteínas principalmente; asociados a los lípidos y las proteínas también se encuentran oligosacáridos.
A. Lípidos; son esencialmente anfipáticos, o sea que sus moléculas poseen un polo hidrófilo y un polo hidrófobo. Los más abundantes (en los glóbulos rojo humano) son los fosfolípidos (55 % del total de los lípidos), el colesterol (25%) y otros lípidos, tales como glucolípidos y ácidos grasos (20 %).

Los lípidos, cuando se encuentran en un medio acuoso, se disponen formando una doble capa, la bicapa lipídica, con las zonas polares (hidrófilas) orientadas hacia el exterior, y las zonas apolares (hidrófobas) protegiéndose mutuamente.

La bicapa lipídica no es una estructura rígida, sino que sus componentes se mueven en ella con libertad, confiriéndole fluidez. Las moléculas de lípidos, más móviles, pueden girar sobre sí mismas (rotación) o intercambiar su posición con otras moléculas de la misma monocapa (difusión lateral). Es poco frecuente el intercambio entre moléculas situadas en monocapas distintas (flip-flop).

El colesterol contribuye grandemente a la fluidez de la bicapa, debido a que con su pequeño tamaño dificulta el establecimiento de interacciones hidrofóbicas entre las colas apolares de los lípidos de membrana.
B. Proteínas; se disponen intercaladas o adosadas a la bicapa de lípidos, y son de diverso tamaño y naturaleza. Por su afinidad a los lípidos de la membrana hay dos tipos:
a) Proteínas integrales o intrínsecas; están internamente asociadas a los lípidos y son difíciles de separar de la bicapa. Constituyen aproximadamente el 70% de las proteínas de membrana y son insolubles en agua (hidrófobas).
b) Proteínas periféricas o extrínsecas; Están débilmente asociadas a los lípidos, se separan con facilidad; son hidrosolubles y representan el 30% restante.

Por su colocación en la membrana, se distinguen los siguientes tipos:
a) Proteínas transmembranarias; atraviesan completamente la membrana.
b) Proteínas de hemimembrana; su posición sólo abarca la mitad de la bicapa.
c) Proteínas adosadas; colocadas por fuera de la bicapa y unidas a proteínas transmembrana o a lípidos.

C. Oligosacáridos; Se asocian a lípidos (glucolípidos) o a proteínas (glucoproteínas).

4.1.2. Estructura de la membrana plasmática
El modelo estructural de membrana más aceptado en la actualidad es el propuesto por Singer y Nicholson y que recibe el nombre de modelo del mosaico fluido. Según éste, todas las membranas celulares responden a un esquema arquitectónico común constituido, básicamente, por una bicapa lipídica a la que se unen los otros componentes de las membranas, es decir: proteínas y glúcidos.
La disposición en bicapa de los lípidos se debe al carácter bipolar que poseen estos constituyentes de la membrana (fosfolípidos, colesterol, glicolípidos, etc.), en un medio acuoso, estos lípidos se disponen enfrentando sus partes hidrófobas dejando sus cabezas hidrófilas en contacto con el medio acuoso intra y extracelular.
La situación de las proteínas (integrales o periféricas) en la membrana la determina su afinidad por el agua o los lípidos. Los azúcares (oligosacáridos), asociados a proteínas (glucoproteínas) o lípidos (glucolípidos), se sitúan en la cara extracelular de la membrana. A esta cubierta se la llama glucocálix o cubierta celular.
Según lo expuesto, el modelo de mosaico fluido de membrana sostiene:
a) Los lípidos y proteínas que forman la membrana plasmática constituyen un mosaico molecular. Están dispuestas unas junto a otras como las piezas de un mosaico.
b) Los lípidos y proteínas pueden desplazarse en el plano de la bicapa lipídica: las membranas son fluidas.
c) Las membranas son asimétricas en cuanto a la disposición de sus componentes moleculares. La asimetría se debe a que la presencia de oligosacáridos está restringida a la superficie de la cara externa y, además, la distribución de los lípidos en una y otra monocapa no es simétrica.

4.1.3. Funciones de la membrana plasmática
a) Frontera física entre dos medios; esto permite no sólo la separación del interior de la célula con respecto al medio exterior (intra y extracelular), sino también la formación de compartimentos en el interior de la célula eucariótica.
b) Facilita que ocurran, de manera simultánea, pero sin mezclarse, una gran diversidad de reacciones químicas en sus diferentes orgánulos.
c) La bicapa lipídica es una eficaz barrera para evitar el paso de sustancias hidrófilas (se evita la pérdida de sustancias intracelulares).
d) Asegura el intercambio y transferencia de sustancias e información con el exterior y con otras células. La naturaleza lipídica de la membrana determina el tipo de sustancias que pueden atravesarla y, además, las proteínas que la forman, pueden intervenir de manera activa facilitando o impidiendo el transporte de esas sustancias.
e) Factores de reconocimiento celular; conforman la “identidad antigénica” de cada individuo debido a que las proteínas específicas de la membrana celular constituyen una combinación única en cada individuo, que permite ser reconocida por las defensas inmunitarias.
f) Receptores hormonales y de otras informaciones; control del flujo de información entre las células y el medio, reciben la información que llega del medio, gracia a la existencia de a) receptores específicos de neurotransmisores y de hormonas, y b) el potencial de membrana responsable de la sensibilidad celular.
g) Desempeñar funciones especiales gracias a las diferenciaciones que presentan algunas: invaginaciones (aumento de la superficie de intercambio), desmosomas (zonas de unión con otras células), etc.
4.1.4. Transporte de sustancias a través de la membrana plasmática
Las membranas son barreras de permeabilidad muy selectiva. Los mecanismos que utilizan las células para permitir el paso de sustancias varían en función de que se trate de moléculas pequeñas, que puedan atravesarla, o de moléculas más grandes, que deban ser englobadas y posteriormente liberadas por la propia membrana:
TRANSPORTE DE MOLÉCULAS PEQUEÑAS; la permeabilidad de las membranas celulares es altamente selectiva. El paso a su través de moléculas e iones está controlado por mecanismos de transporte específicos. Existen dos mecanismos básicos de transporte: el activo y el pasivo.
1. Transporte pasivo; se realiza sin consumo de energía y a favor de un gradiente, ya que la sustancia pasa debido a que hay una diferencia de concentración o de carga eléctrica (o de ambos, electroquímico). Distinguimos:
a) Difusión simple; las sustancias solubles (apolares) pueden atravesar la membrana disueltas en la bicapa lipídica. El agua (y pequeñas moléculas arrastradas por ellas) atraviesa la membrana por ósmosis, lo hace a través de canales acuosos formados por proteínas transmembranarias, desde el lado de menor concentración salina al de mayor.
b) Difusión facilitada; las sustancias polares (azúcares, aminoácidos), para pasar requieren la presencia de proteínas transportadoras (permeasas), a las que se une de manera específica la molécula a transportar y son liberadas de nuevo en el otro lado de la membrana.
La difusión simple y la facilitada se realizan a favor de un gradiente (de concentración o químico, eléctrico, o electroquímico) con lo que no requiere aporte de energía para realizarse.
2. Transporte activo; se realiza en contra de un gradiente (es decir, de la zona más diluida a la más concentrada), se requieren también proteínas transportadoras específicas y un aporte de energía (para realizar el “bombeo”), que se traduce en un consumo de ATP.
Un ejemplo es la bomba de sodio-potasio, que mantiene el potencial electroquímico a ambos lados de la membrana de las células animales (bombea Na+ hacia el exterior de la célula y K+ hacia el interior).
TRANSPORTE DE MACROMOLÉCULAS;
La célula dispone de mecanismos que permiten incorporar o expulsar compuestos de mayor tamaño, por medio de deformaciones de la membrana.
1. Endocitosis; incorpora partículas mediante una invaginación de la membrana en la que quedan incluidas. Luego, la invaginación se estrangula y forma una vesícula en el interior. Se distinguen dos tipos:
a) Pinocitosis; cuando el material captado es líquido (partículas disueltas).
b) Fagocitosis; capta partículas sólidas de mayor tamaño, y se forman vacuolas digestivas. Sólo la realizan células especializadas (por ejemplo, leucocitos).

2. Exocitosis; proceso opuesto al anterior. Se expulsan sustancias contenidas en una vesícula, al unirse ésta a la membrana plasmática y abrirse al exterior.

3. Trancitosis; proceso por el que las sustancias que penetran en la célula por endocitosis tras recorrer la totalidad del citoplasma salen al exterior por un proceso de exocitosis. Ejemplo: transporte de nutrientes por las células de las vellosidades intestinales.
4.1.5. Diferenciaciones de la membrana plasmática
Dependiendo de la función que la célula desempeñe, su membrana plasmática puede presentar diferentes especializaciones:
a) Microvellosidades; son evaginaciones que aumentan la superficie de intercambio. Por ejemplo, en las células intestinales, en las que las moléculas digeridas en el interior del tubo digestivo deben pasar al torrente circulatorio.


b) Invaginaciones; Profundos entrantes con finalidad semejante. Por ejemplo, en las células de los túbulos contorneados de las nefronas (riñón), para la reabsorción de líquido y sales.
c) Uniones intercelulares; para mantener adheridas y comunicadas células vecinas.


4.2 La pared celular vegetal
La pared celular es una forma especializada de matriz extracelular (segregada por la célula y excreta al exterior de la membrana plasmática), que se encuentra adosada a la membrana plasmática de las células vegetales, y que se caracteriza por su alto contenido en celulosa, lo que la hace ser gruesa, rígida y organizada.
4.2.1. Composición química de la pared vegetal
Está formada principalmente por celulosa (homopolisacárido que se origina por la unión β (1→4) de la D-glucosa), pero también por: hemicelulosa, pectinas, sales minerales y agua.
4.2.2. Estructura de la pared vegetal
Está constituida por tres capas, cada una con distinta composición y características. Desde fuera hacia dentro son:
1. Lámina media; es la capa más externa y es común a las dos células adyacentes. Es delgada y flexible, y está compuesta principalmente por pectina. Se encarga de mantener unidas las distintas células en los tejidos vegetales.
2. Pared primaria; capa relativamente delgada y semirrígida, típica de las células jóvenes, recién divididas (plantas en crecimiento). Está formada por celulosa con una abundante matriz hemicelulósica.
3. Pared secundaria; capa muy gruesa formada por tres subcapas de celulosa, en cada una de las cuales las fibras de celulosa se disponen con distinta orientación, lo cual le da a la pared una gran rigidez y resistencia. La pared secundaria sólo se presenta en células maduras o ya muertas. Precisamente el grosor de la capa de celulosa hace que el citoplasma se vaya "asfixiando", y la célula acabe por morir.
4.2.2. Funciones de la pared vegetal
a) Constituyen un exoesqueleto que protege a la célula, le da forma y le confiere resistencia, pero sin impedir su crecimiento.
b) Es la responsable de que la planta se mantenga erguida.
c) Impide que la célula se rompa, ya que interviene activamente en el mantenimiento de la presión osmótica intracelular.
d) Permite la comunicación entre células adyacentes y con el exterior, para el intercambio de nutrientes y de información. Existen unos orificios que atraviesan la pared llamados punteaduras que se sitúan al mismo nivel en células vecinas. Estas punteaduras son atravesadas por puentes citoplasmáticos o plasmodesmos, que son prolongaciones del retículo endoplasmático.
5. El hialoplasma o citosol
El citoplasma es la región comprendida entre la membrana plasmática y la envoltura nuclear, y está constituida por el hialoplasma o "jugo celular" y los orgánulos citoplasmáticos, mantenidos e interconectados por una red de filamentos y túbulos que forman el citoesqueleto o esqueleto celular.
El hialoplasma o citosol, es el medio interno de la célula en el que se encuentran los orgánulos celulares y el núcleo. Está limitado por distintas membranas, la membrana plasmática, la membrana nuclear y las membranas que envuelven los diferentes orgánulos.
Representa entre el 50 y el 80 % del volumen celular, es un medio acuoso (contiene de un 70-80% de agua) en el cual están disueltas gran cantidad de moléculas formando una disolución coloidal (las moléculas forman micelas). Estas moléculas son prótidos, lípidos, glúcidos, ácidos nucleicos, sales minerales e iones.

Es un medio líquido de viscosidad variable (estado sol/gel) y algunas de sus principales funciones son: regular el pH intracelular, almacenar sustancias, medio donde se realizan la mayoría de las reacciones metabólicas celulares, emisión de prolongaciones (pseudópodos) mediante movimientos internos (ciclosis), ...
6. El citoesqueleto
Aparece en todas las células eucarióticas (aunque más desarrollado en las células animales que en las de los vegetales y hongos, debido a que estas últimas poseen pared que cumplen funciones de exoesqueleto celular), y está formado por una red de filamentos proteicos. Estos filamentos son los responsables de las formas de las células, de su movimiento y de su organización interna.
Sus principales funciones son:
a) Mantener la estructura celular (forma general de la célula por andamiaje de microtúbulos, resistencia a la tracción por filamentos intermedios, viscosidad próxima a la membrana plasmática por microfilamentos,…).
b) Movimientos celulares (cilios y flagelos con tubulina y otras proteínas, fibras musculares : actina y miosina, …).
c) Reparto de sustancias (vesículas ligadas a microtúbulos, división celular, …).
d) Fijación de sustancias (anclaje de proteína de membrana a fibras de actina, anclaje de orgánulos a fibras de actina, …).



6.1 Principales tipos de filamentos
6.1.1. Filamentos de actina o microfilamentos
Como su nombre indica, los microfilamentos son estructuras filamentosas, que están constituidos por dos cadenas de proteínas globulares, la actina, enrolladas en hélice, y con un diámetro de 5 nm. Sus funciones son:
a) Mantienen la forma celular, pero con elasticidad.
b) Da rigidez y estabilidad a muchas prolongaciones celulares como microvellosidades, etc.
c) Intervienen en el movimiento ameboide, dando soporte a la emisión de pseudópodos.
d) Intervienen (junto con la miosina) en el movimiento contráctil de las células musculares.
e) Intervienen en la formación de vesículas de endo y exocitosis.
f) Cariocinesis, división celular separando las dos células hijas.


6.1.2. Filamentos intermedios
Son fibras proteicas, gruesas y resistentes. Tienen un diámetro de unos 10 nm (intermedio entre el de los microfilamentos y el de los microtúbulos). Aparecen en células o en regiones celulares sometidas a esfuerzos mecánicos. Hay dos tipos principalmente:
a) Neurofilamentos: dan forma a los axones de las neuronas.
b) Tonofilamentos: filamentos de queratina. Aparecen en las uniones intercelulares adherentes (desmosomas).


6.1.3. Microtúbulos
Son filamentos tubulares, huecos, constituidos por monómeros de tubulina, proteína con forma esférica, existen dos tipos la α-tubulina y la β-tubulina, que se asocian para formar dímeros, los cuales a su vez se unen para formar el microtúbulo con 13 hileras de monómeros. Se forman a partir de centrosoma (o del centro organizador de microtúbulos en las células vegetales). Tienen un diámetro de 25 nm.
Las funciones de los microtúbulos son:
a) Forman estructuras estables, como los centriolos, cilios y flagelos.
b) Forman estructuras lábiles, como los microtúbulos del áster y del huso acromático.
c) Desplazamiento de orgánulos y sustancias citoplasmáticas por la célula.


7. Orgánulos sin membrana
7.1 Centrosoma
El centrosoma o centro celular es una estructura, sin membrana, presente en todas las células animales, excepto en las que no se dividen.
Estructura y composición
El centrosoma consta de un cuerpo central, formado por dos centríolos, rodeado por el material pericentríolar, actualmente al conjunto de los dos centríolos se le llama centro organizador de microtúbulos.
Cada uno de los centríolos están formados por microtúbulos estables, dispuestos en forma de cilindro, y constan de nueve grupos de tres túbulos cada uno (triplete), que se mantienen unidos entre sí. Los tres microtúbulos de cada triplete se encuentran íntimamente asociados, y a su vez, los distintos tripletes están enlazados entre sí por determinadas proteínas que sirven de puente.



Centríolo - Centrioles by Enzo Caldeirini on Sketchfab
Los centrosomas se encuentran próximos al núcleo en las células animales (no aparecen en las vegetales), en una célula en interfase constan de tres partes:
a) Diplosoma; es la parte central y consta de dos centríolos situados cerca del núcleo y dispuestos perpendicularmente entre sí, y rodeados de una porción de hialoplasma íntimamente asociada a ellos.
b) Centrosfera; porción de hialoplasma que rodea al diplosoma, se caracteriza por la carencia de estructuras membranosas.
c) Áster; que consiste en una serie de microtúbulos dispuestos en forma radial
Funciones
a) El centrosoma es el centro organizador de los microtúbulos, forman el cuerpo basal de los cilios y flagelos.
b) Forman el huso acromático o huso mitótico que, como veremos al estudiar la mitosis, es un sistema de microtúbulos que van de un polo de la célula al otro y que se encargan de repartir los cromosomas durante la división celular.
7.2 Cilios y flagelos
Son prolongaciones citoplasmáticas dotadas de movimiento (que permiten el desplazamiento de la célula en un medio acuoso) recubiertas por la membrana plasmática.
Semejanzas y diferencias entre cilios y flagelos:
a) Ambos están constituidos por pares de microtúbulos (no por tripletes).
b) Los cilios se encuentran en gran número, mientras que los flagelos son mucho menos numerosos (normalmente uno o dos).
c) Los cilios son de menos tamaño, 5-10 μm y los flagelos de 100-200 μm.
d) El movimiento de los cilios es de batimiento, es decir, se mueven a un lado y a otro, mientras que los flagelos tienen movimiento ondulatorio.
En su estructura se distinguen 4 zonas:
a) Tallo o Axonema: prolongación citoplasmática recubierta por la membrana. Está compuesto por 9 pares o dobletes de microtúbulos (prolongaciones de los microtúbulos A y B de un centríolo situado en la base) externos unidos entre sí. Presenta un par de microtúbulos centrales. Por lo tanto, el axonema posee 9 dobletes periféricos y 2 microtúbulos centrales.
b) Zona de transición: situada a la altura de la membrana plasmática. Consta de 9 dobletes de microtúbulos. En la zona superior de transición situada debajo del tallo o axonema se encuentra un disco de material amorfo o placa basal en cuyas proximidades nacen los dos microtúbulos de la parte central.
c) Corpúsculo basal o cinetosoma: centríolo en el existe un eje tubular central de donde parten 9 láminas radiales que llegan hasta 9 tripletes de microtúbulos (en el corte esta estructura tiene el aspecto de una rueda de carro).
d) Raíces: microtúbulos que conectan al cilio o flagelo con el citoesqueleto.




Funciones
Ambas estructuras (cilios y flagelos) están directamente relacionadas con el movimiento. En el caso de los flagelos su movimiento ondulatorio puede producir un desplazamiento de la célula libre (no fijadas a tejidos). En los cilios, normalmente el movimiento de batimiento tiene como objetivo renovar el líquido extracelular en contacto con la célula, lo cual a su vez suele estar relacionado con procesos de nutrición celular.

7.3 Ribosomas
Los ribosomas son orgánulos celulares globulares sin membrana, sólo visibles con el microscopio electrónico. Químicamente están compuesto por ARNr (ribosomico), proteínas y gran cantidad de agua. Aparecen en todos los tipos de celulas, procariótas y eucariótas que pueden estar libres en el hialoplasma o agrupados, polirribosomas o polisomas mediante un ARNm, ligados al retículo endoplasmático, así como en el interior de las mitocondrias y los cloroplastos.
Estructura
En las procarióticas los ribosomas son de menor tamaño (70 S) que en las eucarióticas (alrededor de 80 S). La “S” (unidades Svedberg) representa una unidad de medida que equivale a 10-3 sg. y mide el tiempo que un ribosoma tarda en sedimentarse en una centrífuga, lo que depende fundamentalmente de su forma y tamaño.

Estructuralmente los ribosomas de las células eucariotas están formados por dos subunidades, una mayor (60 S) y otra menor (40 S). Las dos subunidades se forman en el nucléolo donde se unen sus dos componentes: el ARNr y las proteínas ribosomales. El ARNr se sintetiza en el núcleo, mientras que las proteínas lo hacen en el citoplasma y posteriormente migran hacia el núcleo. Las dos subunidades ribosomales salen al citoplasma por los poros nucleares y es allí donde se unen para formar el ribosoma.



8. Orgánulos con una membrana
8.1 Retículo endoplasmático (R.E.)
Está formado por un conjunto de membranas que limitan cavidades cerradas o cisternas de distintas formas: sáculos aplanados, vesículas globulares o tubos de aspecto sinuoso que delimitan un espacio interno, lumen o espacio cisternal, contenido en el interior del retículo endoplasmático y el espacio citosólico en el exterior del retículo. Estas cavidades se comunican a menudo entre ellas y forman una red (retículo = pequeña red) característica de las células eucariotas.
Sacos del retículo, por lo tanto, dobles membranas, separan el material nuclear del resto del citoplasma formando la membrana nuclear y, por lo tanto, separando el núcleo del citoplasma. Esta doble membrana está atravesada por una serie de poros que permiten el paso de grandes moléculas (ARN, subunidades ribosómicas, ...).
Podemos distinguir dos tipos de retículo:
a) Retículo endoplasmático rugoso (R.E.R.); que posee ribosomas adheridos a la cara de la membrana que da al hialoplasma (espacio citosólico). Se encuentra muy desarrollado en aquellas células que participan activamente en la síntesis de proteínas.
Dentro de sus funciones destacan: la síntesis de proteínas por parte de los ribosomas que pueden ser almacenadas en el lumen del retículo donde maduran sufriendo con frecuencia una glicosilación (se unen a un glucido, glucoproteínas) y son transportadas hacia otros orgánulos (aparato de Golgi, lisosomas), a la membrana celular o las propias membranas del retículo.

b) Retículo endoplasmático liso (R.E.L.); constituido por finos túmulos o canalículos interconectados y cuyas membranas se continúan con las del R.E.R., pero sin llevar adheridos ribosomas.
Sus funciones son: la síntesis de lípidos de membrana (fosfolípidos, colesterol, etc.); síntesis, almacenamiento y transporte de lípidos, y la detoxificación de aquellas sustancias perjudiciales para la célula producidas por su actividad vital o procedente del exterior (insecticidas, herbicidas, medicamentos, etc.). Las toxinas que proceden del medio externo llegan a nuestras células, son transformadas por este orgánulo en moléculas cuya toxicidad se ha visto reducida y que son de esta manera más fácilmente eliminadas. Otras funciones destacables son actuar como un reservorio intracelular de Ca2+ o defosforilar la glucosa-6P para que pueda salir por la membrana celular.


8.2 Aparato de Golgi
El aparato de Golgi es un orgánulo membranoso que forma un sistema de cavidades aplanadas dispuestas ordenadamente. Cada una de estas cavidades recibe el nombre de sáculo. El conjunto de sáculos apilados recibe el nombre de dictiosoma. Además, pueden observarse toda una serie de vesículas más o menos esféricas a ambos lados y entre los sáculos relacionadas con el transporte de proteínas y lípidos desde y hacia el aparato de Golgi.

El conjunto de todos los dictiosomas y vesículas celulares constituye el aparato o complejo de Golgi. Se encuentra más desarrollado cuanto mayor es la actividad celular y en las células secretoras.
El aparato de Golgi suele situarse, en la célula animal, cerca del centrosoma pues su funcionamiento depende en gran medida del sistema de microtúbulos de la célula. En las células vegetales, que no tienen centrosoma, no se aprecian dictiosomas como tales y las vesículas del aparato de Golgi están más dispersas.
El aparato de Golgi es una estructura variable y en constante renovación; durante la mitosis o cuando el tráfico de sustancias en la célula se detiene prácticamente desaparece.
El aparato de Golgi es un orgánulo polarizado. En un dictiosoma se diferencian dos caras diferentes, una cara cis o de entrada y una cara trans o de salida. La cara cis está relacionada con el retículo endoplasmático y de la cara trans surgen diferentes vesículas de transporte que se dirigen a sus destinos finales: la membrana plasmática, los lisosomas y las vesículas de secreción.

Funciones:
a) Embalaje y secreción; las sustancias fabricadas en el R. E. para ser secretadas se incorporan a la cara “cis” o de formación del aparato de Golgi y se desplazan progresivamente hacia la cara “trans” o de maduración, desde donde se incorporan a las vesículas de secreción. Las vesículas se adosan a la membrana y por un proceso de exocitosis vierten su contenido al medio extracelular.

b) Reciclaje de la membrana plasmática; el proceso anterior genera un reciclaje permanente de la membrana compensado por los mecanismos de endocitosis. Entre los productos secretados tienen especial interés los que forman la matriz extracelular y, en los vegetales, la pared celular vegetal.
c) Formación de lisosomas y vacuolas; no todos los productos empaquetados por el aparato de Golgi tienen como destino el exterior de la célula. Por ejemplo, algunas vesículas, que contienen abundantes enzimas hidrolíticos, se transforman en lisosomas y permanecen en el citoplasma celular. En las células vegetales originan las vacuolas.
d) Glicosilación; los compuestos que vienen del R.E. experimentan modificaciones, glucolisaciones, por la adición de oligosacáridos, que se unen a proteínas (glucolisación de proteínas: glucoproteínas de membrana, anticuerpos) y a lípidos (glucolisación de lípidos: glucolípidos de membrana).
8.3 Lisosomas
Los lisosomas son orgánulos celulares globulares, aunque presentan formas muy diversas. Se originan a partir del aparato de Golgi, de la misma manera que se originan las vesículas de secreción.

Constituyen el aparato digestivo de la célula. Contienen un ambiente ácido y enzimas hidrolíticas (fosfatasa ácida, proteasa,...) que catalizan la hidrólisis o digestión de las macromoléculas; la membrana que limita a cada lisosoma impide que la célula sea digerida por estos enzimas líticos al estar muy glicosilada. Tiene una bomba de protones que mantiene el pH ácido del interior y proteínas de transporte que permiten que los productos de la digestión pasen al citosol.
Se encuentran en las células eucariotas tanto animales como vegetales, la estructura de estos orgánulos es muy distinta de un tipo celular a otro e incluso en una misma célula. Este polimorfismo hace que no sea posible caracterizar a los lisosomas bajo criterios estrictamente estructurales y tan sólo la demostración de actividades líticas permite poner de manifiesto la naturaleza lisosómica de una inclusión celular. A pesar de estas diferencias, los lisosomas se clasifican en:
a) Lisosomas primarios; recién formados a partir del aparato de Golgi. Tienen una forma ovalada o redondeada y solo contienen enzimas hidrolíticos pues todavía no han intervenido en ningún proceso de digestión celular.
b) Lisosomas secundarios; de forma variable (depende de la naturaleza de los sustratos y del estado de digestión en el que se encuentren), están implicados en algún proceso de digestión. Se forman a partir de la unión de los lisosomas primarios con endosomas que contienen sustratos procedentes del medio externo (vacuolas heterofágicas o digestivas) o interno (vacuolas autofágicas).
c) Cuerpos residuales; contienen material que ya no puede ser degradado y quedan almacenados en el interior celular o se funden con la membrana plasmática expulsando estos materiales al exterior (exocitosis).
8.3.1 Funciones:
A.- Digestión intracelular
a) Vacuolas heterofágicas o digestivas (heterofagia); trabajan con sustratos procedentes del medio externo y su finalidad es alimenticia y defensiva.
Las partículas procedentes del medio externo penetran en la célula, mediante un proceso de invaginación de la membrana celular (pino o fagocitosis), que se separa de ésta, formándose una vacuola alimenticia o vacuola de endocitosis. Los lisosomas primarios se unen a esta vacuola, dando lugar a un orgánulo único, la vacuola digestiva o lisosoma secundario, que contiene no sólo enzimas hidrolíticos, sino también moléculas que va a digerir, por lo que se considera a esta vacuola el estómago celular.
Estas moléculas, una vez que son digeridas, pasan al citoplasma celular. Sin embargo, no todas las sustancias son digeridas y por tanto no pueden ser aprovechadas por la célula. A estas partículas no digeridas se las llama productos residuales, y, al lisosoma, cuerpo residual siendo almacenados en el interior celular o expulsados al exterior de la célula (exocitosis).
b) Vacuolas autofágicas (autofagia); si el sustrato lo constituyen componentes de la propia célula. Este mecanismo interviene en los procesos de desarrollo, aprovechando aquellos orgánulos o materiales obsoletos, y asegura la nutrición en condiciones desfavorables.

B.- Digestión extracelular
En algunas ocasiones, los lisosomas vierten su contenido hidrolítico al exterior de la célula, donde tiene lugar la digestión (digestión extracelular).
8.4 Peroxisomas
Los peroxisomas son orgánulos delimitados por membrana que contienen enzimas oxidativas, en ocasiones en tal cantidad que es posible observar inclusiones cristalinas en su interior. Deben su nombre a que las primeras enzimas que se descubrieron en su interior fueron las peroxidasas, pero pueden encontrarse en su interior más de 50 enzimas encargadas de catalizar diversas reacciones, que varían dependiendo del tipo celular y del estado fisiológico de la célula.

Están especializados en llevar a cabo reacciones que utilizan el O2 generando peróxido de hidrógeno (H2O2), que por ser un agente oxidante muy tóxico, es utilizado a continuación por la catalasa para realizar reacciones oxidativas útiles.
R-H2 + O2 ----> R + H2O2 (muy tóxico)
H2O2 + catalasa ----> 2 H2O + O2

Su origen ha sido discutido porque sus proteínas se forman en el citosol pero se ha observado que pueden formarse a partir del R.E. Por otra parte tienen la capacidad de dividirse de forma autónoma, como ocurre en la mitosis. Actualmente la tendencia más aceptada piensa que son la evolución de un orgánulo primitivo pre-mitocondrias responsable de todo el metabolismo oxidativo; con la aparición de las mitocondrias que pueden hacer este metabolismo y aprovecharlo para producir grandes cantidades de ATP la importancia de los peroxisomas paso a ser secundaria.
Funciones:
a) Intervienen en reacciones de detoxificación. Los grandes peroxisomas de las células hepáticas y renales detoxifican diversas moléculas que entran en circulación. Por ejemplo, casi la mitad del etanol que bebemos es oxidado a acetaldehido por acción de la catalasa.
b) Llevan a cabo reacciones oxidativas de degradación de ácidos grasos y aminoácidos, generando H2O2. Estas reacciones no proporcionan a la célula energía útil en forma de ATP, pero producen calor, que en ocasiones tiene importancia fisiológica.
c) En las plantas y los hongos, produce la ß-oxidación de los ácidos grasos (en los animales también se realiza en las mitocondrias).
d) En las plantas realiza la fotorrespiración (oxidación de productos residuales de la fotosíntesis que consume O2 y libera CO2.
8.5 Glioxisomas
Orgánulos exclusivos de las células vegetales y de los hongos filamentosos.
Son parecidos a los peroxisomas (derivan y pueden convertirse en ellos), y se encargan, durante la germinación, de transformar los lípidos almacenados en la semilla en glúcidos necesarios para el crecimiento del embrión hasta que la plántula pueda realizar la fotosíntesis.
Para poder hacer esto (imposible en las células animales) poseen enzimas del ciclo del ácido glioxílico que es una variante del ciclo de Krebs de las mitocondrias que permite sintetizar azúcares a partir de grasas. Es indispensable en semillas en germinación.

8.6 Vacuolas
Las vacuolas son vesículas de forma globular y tamaño variable destinadas a almacenar todo tipo de sustancias. Se forman a partir del R.E., del aparato de Golgi o de la membrana plasmática (endocitosis).
El conjunto de vacuolas de una célula se denomina vacuoma. Las células vegetales presentan vacuolas de gran tamaño, que a veces desplazan a la periferia todo el citoplasma celular y el núcleo, y son escasas en número. Esta vacuola recibe el nombre de vacuola central. En cambio, las células animales poseen numerosas vacuolas de pequeño tamaño, habitualmente denominadas vesículas.

Funciones:
a) Almacenar sustancias de reserva: lípidos, ácidos grasos, sales minerales, proteínas solubles, proteínas precipitadas (granos de aleurona), ...
b) Almacenar sustancias especiales: taninos, pigmentos, enzimas hidrolíticos (cuya actividad, dada su función de reserva, se encuentra inhibida en tanto no sean necesarias),...
c) Servir de vehículo de transporte de sustancias entre orgánulos citoplasmáticos (vesículas de transición, de secreción, etc.) y entre el exterior e interior de la célula (vacuolas o vesículas de endocitosis).
d) Regulan la presión osmótica de la célula; las vacuolas pueden llenarse o vaciarse de agua para mantener constante la concentración del citoplasma.
e) Vacuolas pulsátiles; eliminan el exceso de agua citoplasmática en células de organismos dulceacuícolas (que son hipertónicos respecto al medio).
9. Orgánulos con doble membrana: Mitocondrias y plastos
9.1 Mitocondrias
Las mitocondrias son orgánulos presentes en el citoplasma de todas las células eucarióticas (aeróbicas). Su forma se asemeja a un cilindro alargado y son difíciles de observar al microscopio óptico, al que aparecen como palitos o bastoncitos alargados.
Lo más frecuente es que las mitocondrias estén dispersas en el hialoplasma, y su número depende del tamaño de la célula: unas pocas en las levaduras y de 1000 a 2000 en una célula hepática. Son las encargadas de la obtención de la energía mediante la respiración celular, proceso de oxidación en el que intervienen las ATP sintetasas. La energía obtenida se guarda en forma de ATP.
Ultraestructura
Podemos distinguir las siguientes partes (de fuera hacia dentro):
a) Membrana mitocondrial externa; membrana continua de 60 Å de espesor que separa el interior de la mitocondria del hialoplasma. Su estructura es la misma que la del resto de las membranas celulares y es muy permeable debido a que posee un gran número de proteínas que forma “canales” a través de los que pasan gran cantidad de moléculas.
b) Espacio intermembrana; de composición similar al hialoplasma por la permeabilidad de la membrana externa. Las enzimas que posee le permiten transferir la energía del ATP fabricado en la mitocondria a otros nucleótidos, por ejemplo el AMP (AMP + ATP ▬► 2 ADP). Las moléculas de ADP así formadas pueden atravesar la membrana interna y ser fosforiladas en moléculas de ATP.
c) Membrana mitocondrial interna; posee una gran superficie (unas 5 veces más que la externa) debido a los repliegues que forman las crestas mitocondriales. Es impermeable a gran cantidad de sustancias (iones,...). No contiene colesterol (como la membrana plasmática bacteriana). Contiene un 20% de lípidos y un 80% de proteínas, las cuales se pueden clasificar en tres grupos:
1) Proteínas transportadoras que regulan el paso de metabolitos a través de la membrana interna (cadena transportadora de electrones).
2) Enzimas de la cadena respiratoria, que catalizan las reacciones de oxidorreducción.
3) Complejo enzimático ATP-sintetasa, cataliza la producción de ATP en la matriz, poseen tres subunidades: la esfera F1 o partícula elemental pegada a la cara matricial de la membrana interna (es la parte catalítica del complejo), el pedúnculo F0, que une las esferas a la membrana, y la base hidrófoba integrada en la membrana mitocondrial interna.

d) Matriz mitocondrial; espacio interno que contiene numerosos iones y moléculas solubles, en particular una gran variedad de enzimas para llevar a cabo las rutas metabólicas que tienen lugar en su interior. Además, contiene ADN mitocondrial, ribosomas 70S (mitorribosomas), ARNt, ...


Funciones
a) Oxidaciones respiratorias; cuya finalidad es obtener energía para la célula a partir de materia orgánica. Consisten en la degradación completa de moléculas orgánicas a CO2 para liberar energía y formar ATP. Tienen lugar en dos series de procesos: La β-oxidación de los ácidos grasos y el ciclo de Krebs; y la cadena de transporte de electrones, que suceden, respectivamente, en la matriz y en la membrana mitocondrial interna.
b) Producción de metabolitos precursoras para la biosíntesis de macromoléculas que se sintetizan en el hialoplasma.
c) Síntesis de proteínas mitocondriales, ya que, poseen la maquinaria y la información necesaria para ello.

9.1 Plastos
Los plastos, o plastidios, son un conjunto de orgánulos exclusivos de células vegetales que tienen un origen común. Poseen, al igual que las mitocondrias, información genética propia, ribosomas y una doble membrana. Hay tres tipos de plastos, aunque su estructura es muy semejante:
a) Leucoplastos (leuco = blanco); aparecen en partes no verdes del vegetal - las subterráneas, por ejemplo- entre ellos destacan los amiloplastos que acumulan almidón en los tejidos de reservas.
b) Cromoplastos (cromo = color); dan color rojo, anaranjado o amarillo a muchos frutos, flores y otros órganos. Acumulan pigmentos fotosintéticos secundarios (xantofilas y carotenos) y reservas nutritivas.
c) Cloroplastos (cloro = verde); son de color verde debido a la clorofila que contienen. Se encargan de realizar la fotosíntesis, captando energía luminosa.
9.1.1. Cloroplastos
Los cloroplastos se localizan en las células vegetales fotosintéticas y, en los vegetales superiores, tienen forma lenticular (de lenteja). Su número suele ser de unos 40 por célula. Son de color verde debido a su elevado contenido en clorofila.
Ultraestructura
La observación al microscopio electrónico de un cloroplasto de una célula de vegetal superior nos revela que cada cloroplasto está rodeado de una doble membrana (externa e interna), la membrana externa separa el cloroplasto del hialoplasma, y la membrana interna, que delimita un estroma. Entre ambas existe un espacio, el espacio intermembrana.
La membrana interna cloroplástica (M.I.C.), a diferencia de lo que ocurre en las mitocondrias, carece de crestas y encierra un gran espacio central, el estroma (que contiene ribosomas, enzimas, ADN y ARNt , gránulos de almidón y gotas de lípidos), en el que se bañan un tercer tipo de membrana, la membrana tilacoidal, que constituyen las paredes de unas pequeñas vesículas discoidales aplanadas (a modo de largos sacos cerrados), los tilacoides.
En las membranas de los tilacoides se encuentran los pigmentos fotosintéticos, fundamentalmente clorofila y carotenoides. Los tilacoides están comunicados entre sí y encierran un tercer compartimento el espacio tilacoidal (o intratilacoidal). Existen unos tilacoides más alargados que a veces comunican con la M.I.C. y que son llamados lamelas o "tilacoides del estroma".
Los tilacoides no se distribuyen de manera uniforme por el estroma y en ocasiones forman pequeños grupos apilados (a modo de pilas de monedas) llamados grana (plural de grano en latín) debido al aspecto que presentan al observarlos con el microscopio. Normalmente los tilacoides se disponen paralelamente al eje mayor del cloroplasto.



Funciones
Una de ellas es la síntesis de proteínas cloroplásticas, ya que, como hemos visto, poseen la maquinaria y la información necesaria para ello.
Otra función es servir como un lugar de almacenamiento temporal de almidón (su acumulación suele formar gránulos, con frecuencia de gran tamaño, que suelen poder observarse con facilidad).

Pero la su principal función es llevar a cabo la fotosíntesis, que consiste en la formación de materia orgánica a partir de compuestos inorgánicos (H2O, CO2, NO3=, SO4=, PO43-...), que reducen gracias a la energía luminosa captada por los pigmentos fotosintéticos.

En los tilacoides se encuentran los pigmentos captadores de luz y la cadena transportadora de electrones (o cadena fotosintética), donde el ADP sé fosforila y pasa a ATP. Al ser la luz, digamos, el motor de este proceso, se le da el nombre de fotofosforilación. En los tilacoides se produce también la reducción del NADP+, que pasa a NADPH + H+. Ambos procesos constituyen lo que se conoce como fase lumínica de la fotosíntesis.
La energía acumulada, tanto en el ATP como en el NADPH + H+, será utilizada en la fase oscura para la fijación del CO2 a moléculas orgánicas. Esta fase oscura de la fotosíntesis tiene lugar en el estroma del cloroplasto.
Como ocurre en la mitocondria, en los cloroplastos se encuentran también esferas de ATP-sintetasa (o ATPasa), situadas en la membrana interna del cloroplasto que mira hacia el estroma y en la membrana de los tilacoides. La ATPasa cataliza la reacción de formación de ATP .


10. El núcleo celular
El núcleo es la estructura más destacada de la célula eucarionte, tanto por su morfología como por sus funciones. Su tamaño es variable, al igual que su ubicación siendo en la mayoría de los tipos celulares central.
El núcleo tiene tres funciones primarias (todas ellas relacionadas con su contenido de ADN):
a) Almacenar la información genética en el ADN.
b) Recuperar la información almacenada en el ADN en la forma de ARN.
c) Ejecutar, dirigir y regular las actividades citoplasmáticas, a través del producto de la expresión de los genes: las proteínas.
En el núcleo se localizan los procesos a través de lo cuales se llevan a cabo dichas funciones. Estos procesos son:
a) La duplicación del ADN y su ensamblado con proteínas (histonas) para formar la cromatina.
b) La transcripción de los genes a ARN y el procesamiento de éstos a sus formas maduras, muchas de las cuales son transportadas al citoplasma para su traducción.
c) La regulación de la expresión genética.
10.1 Estructura del núcleo
El núcleo está rodeado por la envoltura nuclear, una doble membrana interrumpida por numerosos poros nucleares. Los poros actúan como una compuerta selectiva a través de la cual ciertas proteínas ingresan desde el citoplasma, como también permiten la salida de los distintos ARN y sus proteínas asociadas.
La envoltura nuclear es sostenida desde el exterior por una red de filamentos intermedios dependientes del citoesqueleto, mientras que la lámina nuclear, la cual se localiza adyacente a la superficie interna de la envoltura nuclear, provee soporte interno.
El núcleo también tiene un nucleoplasma, en el cual están disueltos sus solutos y un esqueleto filamentoso, la matriz nuclear la cual provee soporte a los cromosomas y a los grandes complejos proteicos que intervienen en la replicación y transcripción del ADN.
Los cromosomas aparecen ocupando lugares específicos. Los genes que codifican productos relacionados, aunque estén localizados en diferentes cromosomas, pueden estar ubicados próximos en el núcleo interfásico. Por ejemplo, los cromosomas humanos 13, 14, 15, 21 y 22 poseen un gran número de genes que codifican para ARNr. Dichos cromosomas están agrupados de tal forma que los genes de los ARNr están todos juntos y confinados en el nucléolo, el lugar donde se sintetizan, procesan y ensamblan los ARNr. Esta separación física asegura que los ARNr puedan ser eficientemente ensamblados dentro de las subunidades ribosomales.
En el núcleo, los genes transcripcionalmente activos tienden a estar separados de los inactivos. Los activos se encuentran ubicados centralmente, mientras que los silentes están confinados próximos a la envoltura nuclear.
Tan pronto como las células entran en mitosis o meiosis, los fragmentos de la matriz nuclear dirigen la condensación de los cromosomas, constituyéndose en la parte central de los mismos.


10.2 La envoltura nuclear
La envoltura está formada por dos membranas concéntricas interrumpidas por poros nucleares y por la lámina nuclear.
Las membranas delimitan un espacio de 10 a 50 nm, el espacio o cisterna perinuclear. La membrana externa en contacto con el citoplasma tiene ribosomas adheridos, que sintetizan las proteínas que se vuelcan al espacio perinuclear. El espacio perinuclear se continua con el R.E.R.
La envoltura nuclear es un derivado del sistema de endomembranas, siendo esto evidente al inicio de la división celular, cuando la envoltura se desorganiza y pasa a formar parte del sistema de cisternas y vesículas del retículo endoplásmico.
La aparición de la envoltura nuclear permitió que los eucariontes aislaran los procesos genéticos principales, como la autoduplicación del ADN o la síntesis de ARN. Además esto posibilitó que el ARNm se modifique dentro del núcleo antes de ser traducido en los ribosomas. Estas modificaciones no ocurren en los procariontes, ya que a medida que la ARN polimerasa sintetiza el ARN, simultáneamente el extremo 5’ se une al ribosoma y comienza la traducción.



10.3 Cromatina y cromosomas
El núcleo contiene los cromosomas de la célula. Cada cromosoma consiste en una molécula única de ADN con una cantidad equivalente de proteínas. Colectivamente, el ADN con sus proteínas asociadas se denomina cromatina.
La mayor parte de las proteínas de la cromatina consisten en copias múltiples de cinco clases de histonas. Estas proteínas básicas son ricas en residuos de arginina y lisina cargados positivamente. Por esta razón se unen estrechamente con los grupos fosfatos (cargados negativamente) del ADN.
La cromatina también contiene pequeñas cantidades de una amplia variedad de proteínas no histónica. La mayoría de ellas son factores de transcripción (por ejemplo, el receptor esteroide), siendo su asociación con el ADN pasajera. Estos factores regulan que parte del ADN será transcripta en ARN.
10.3.1 Niveles de organización de la cromatina
La observación a través del microscopio óptico de un núcleo interfásico, nos permite distinguir dos tipos de cromatina. La eucromatina o cromatina laxa, de localización central, y la heterocromatina o cromatina densa, en la periferia del núcleo. La heterocromatina representa aproximadamente el 10% del total de cromatina y es considerada transcripcionalmente inactiva.

La eucromatina se encontraría al menos en dos estados: la eucromatina accesible, que representa alrededor del 10%, donde se encuentran los genes que se están transcribiendo y la eucromatina poco accesible, más condensada (pero menos que la heterocromatina, donde están los genes que la célula no está transcribiendo.
La forma menos condensada de la cromatina tiene la apariencia de un collar de perlas. Las perlas son los nucleosomas, las unidades básicas de enrollamiento de la cromatina. Los nucleosomas están formados por un centro o "core" de histonas. Dicho centro posee dos copias de cada una de las siguientes histonas: H2A; H2B; H3 y H4.
Alrededor del centro de histonas, 146 pares de bases del ADN se enrollan en dos vueltas. La unión de las histonas al ADN no depende de una secuencia particular de nucleótidos, sino de la secuencia de aminoácidos de la histona. Las histonas son unas de las moléculas más conservadas durante el transcurso de la evolución (lo cuál es reflejo de su importancia).
Alrededor de 60 pares de bases de ADN unen un nucleosoma con el próximo. Cada región de unión es el ADN espaciador. La quinta histona, la H1, conecta a los nucleosomas y actúa como una banda de goma, manteniéndolos juntos dentro de una misma cuerda enrollada. Esta estructura se conoce como fibra de 10 nm o collar de cuentas, siendo el primer grado del empaquetamiento de la cromatina.

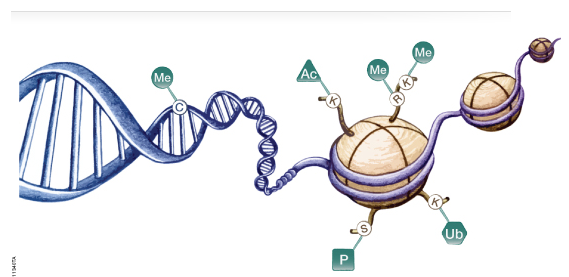

Los nucleosomas se organizan, a su vez, en fibras de 30 nm (solenoide), segundo grado del empaquetamiento de la cromatina girando a manera de resorte alrededor de un eje virtual. Esta estructura es mantenida por la interacción de las H1 de nucleosomas cercanos.


En el siguiente nivel de empaquetamiento, las fibras de 30 nm se organizan en una serie de bucles o asas superenrolladas, tercer grado de empaquetamiento de la cromatina. Estos bucles se estabilizan gracias a la interacción con las proteínas de la matriz nuclear o andamiaje nuclear.
Cada bucle de cromatina representa un dominio funcional o unidad de replicación (Fig. 10.10e). Estos dominios contienen alrededor de 100.000 pares de bases, extensión de ADN suficiente para acomodar varios genes de tamaño promedio. Algunos genes, sin embargo, pueden abarcar varios dominios adyacentes de un cromosoma. Cada cromosoma puede tener cien o más dominios. Durante la profase, los cromosomas aparecen en forma más condensada, alcanzando la cromatina su mayor nivel de condensación en metafase. La organización de los cromosomas envuelve la fosforilación de la H1 y otras proteínas, lo cual causa el plegamiento y empaquetamiento aún más compacto de la cromatina. El andamiaje o matriz nuclear se convierte en el centro de la estructura del cromosoma, y como la compactación continúa, éste se pliega modo de acordeón (cuarto grado de empaquetamiento de la cromatina).


El quinto grado de empaquetamiento de la cromatina ya sería el cromosoma metacéntrico (máxima condensación posible). El grado de condensación de los dominios de cromatina se mantiene principalmente debido a la asociación con la matriz nuclear y a proteínas asociadas como la topoisomerasa II o girasa, encargada de controlar el grado de superenrollamiento del ADN . La unión entre la cromatina y la matriz se da a nivel de zonas altamente conservadas, denominadas secuencias SAR o MAR.


Resumen del empaquetamiento:


10.3.2 El cromosoma eucariota
Cada cromosoma eucariota consiste en una molécula simple de ADN de alrededor de 150 millones de pares de nucleótidos.
La molécula de ADN en el cromosoma eucariota es lineal, por lo tanto posee dos extremos (en contraste con el cromosoma bacteriano que es circular).
La molécula de ADN de un cromosoma típico eucariota contiene un conjunto lineal de genes que codifican para ARN y proteínas interrumpido por muchas secuencias de ADN no codificante.
El ADN no codificante incluye:
a) Secuencias de aproximadamente 170 nucleótidos de ADN satélite, repetidas miles de veces, que corresponden al centrómero. El centrómero es muy importante en la separación de las cromátidas en la mitosis. Dependiendo de su posición distinguimos cuatro tipos de cromosomas:

b) Secuencias repetitivas en los extremos del cromosoma llamadas telómeros. Estos extremos son muy importantes para proteger al cromosoma de las nucleasas y que el cromosoma interaccione con la envoltura nuclear.
c) Múltiples secuencias señalizadoras altamente conservadas, denominadas origen de replicación (ORI), necesarias para que se realice la duplicación del ADN en un tiempo breve.

Todas las especies de seres vivos diploides tienen un número fijo de pares de cromosomas homólogos que se representa con el símbolo 2n. Los seres humanos tenemos 23 pares de cromosomas homólogos luego nuestro 2n = 46.
El cariotipo es una representación gráfica o fotográfica de los cromosomas presentes en el núcleo de una sola célula somática de un individuo ordenados. Cada miembro del par de cromosomas homólogos proviene de cada uno de los padres del individuo cuyas células examinamos. En los seres humanos hay 22 pares de autosomas (pares de cromosomas iguales) y un par de heterosomas o cromosomas sexuales (el 23).
El par 23 de las mujeres tiene dos cromosomas XX y el de los hombres un cromosoma X y un cromosoma Y; en el cromosoma Y hay un gen (el gen SRY) que desencadena la formación de los testículos y, por lo tanto, de los caracteres masculinos.

10.4 El nucleolo
En el nucléolo tiene lugar la formación de subunidades ribosómicas, la síntesis y procesamiento de ARNr y actualmente se considera que desempeña un importante papel en la regulación del ciclo celular.
El nucléolo es un aglomerado de fibras de cromatina de distintos cromosomas. En el hombre, los pares 13,14, 15, 21 y 22, aportan sectores de cromatina que forman el nucléolo. Todos estos cromosomas son acrocéntricos y presentan constricciones secundarias denominadas organizadores nucleolares (NOR), donde están los genes que codifican ARNr.
El tamaño del nucleolo varía entre células y en la misma célula según su actividad, pues si bien la velocidad de transcripción puede acelerarse, el ensamblado de las subunidades ribosomales requiere de un tiempo más o menos constante; es por ello que en los nucléolos grandes observamos mayor proporción de componente granular.

11. El ciclo celular
El ciclo celular (también llamado ciclo de división celular) es una secuencia de sucesos que conducen en primer lugar al crecimiento de la célula y posteriormente a la división de la misma en células hijas. Se inicia en el instante en que aparece una nueva célula, descendiente de otra que se ha dividido, y termina en el momento en que dicha célula, por división subsiguiente, origina nuevas células hijas.
El ciclo celular es la base para la reproducción de los organismos. Su función no es solamente originar nuevas células sino asegurar que el proceso se realice en forma debida y con la regulación adecuada (con controles internos para evitar la posible creación de células con múltiples errores). La creación de nuevas células permite al organismo mantenerse en un constante equilibrio, previniendo así aquellos desórdenes que puedan perjudicar su salud (enfermedades congénitas, cáncer, etc.). Los controles internos en la célula son ejecutados por proteínas que no permiten que se presenten situaciones desastrosas (enfermedades) para un ser vivo. Las células que no entrarán en división no se consideran que estén en el ciclo celular.
En rigor, el ciclo celular (la secuencia de sucesos) comprende dos periodos bien nítidos: la interfase (etapas G1, S y G2) y la división celular (etapa M). Esta ultima tiene lugar por mitosis o meiosis.
La interfase es el período comprendido entre divisiones celulares. Es la fase más larga del ciclo celular, ocupando casi el 95% y, como ya hemos dicho, se divide en tres subetapas: G1, S y G2.
El estado o etapa G1, del inglés Growth o Gap1 (Intervalo 1), es la primera fase del ciclo celular, en la que existe crecimiento celular con síntesis de proteínas y de ARN. Es el período que trascurre entre el fin de una mitosis y el inicio de la síntesis de ADN. Tiene una duración de entre 6 y 12 horas, y durante este tiempo la célula duplica su tamaño y masa debido a la continua síntesis de todos sus componentes, como resultado de la expresión de los genes que codifican las proteínas responsables de su fenotipo particular. Al final de esta etapa se distingue un momento de no retorno, llamado punto de restricción o punto R, a partir del cuál ya es imposible detener la sucesión de las fases S, G2 y M.
El estado o etapa S (del inglés Synthesis) es la segunda fase del ciclo, en la que se produce la replicación o síntesis del ADN, como resultado cada cromosoma se duplica y queda formado por dos cromátidas idénticas. Con la duplicación del ADN, el núcleo contiene el doble de proteínas nucleares y de ADN que al principio. Tiene una duración de unos 6-8 horas.
El estado o etapa G2 o Gap2 (Intervalo 2), es el tiempo que transcurre entre la fase S y el inicio de la mitosis (la célula se prepara para mitosis). Tiene una duración entre 3 y 4 horas. Termina cuando la cromatina empieza a condensarse al inicio de la mitosis.


El final de la mitosis da cabida a un nuevo ciclo en G1 o puede que la célula entre en fase G0 que corresponde a un estado de reposo especial característico de algunas células, en el cual puede permanecer por días, meses y a veces años. Las células que se encuentran en el ciclo celular se denominan proliferantes y las que se encuentran en fase G0 se llaman células quiescentes.
Como todo proceso orgánico, el ciclo celular está sujeto a regulación. Ésta es realizada en sitios específicos llamados puntos de control o de chequeo, que pueden frenar o disparar diversos procesos que le permitan a la célula proseguir con su ciclo normal de replicación del material genético, crecimiento y división. La función de la regulación, básicamente es realizada por proteínas específicas conocidas como cinasas (kdc) y ciclinas (ciclinas A ó B).
Hay células que se encuentran permanentemente en el ciclo, como las epiteliales; otras están permanentemente fuera del ciclo, como las neuronas, y otras están fuera del ciclo, pero bajo un estímulo adecuado pueden volver a dividirse, como es el caso de las células hepáticas.
Durante la fase anterior G1, las moléculas de ADN están enrolladas alrededor de las proteínas histonas, formando los fibras de cromatina que constituyen los cromosomas. Durante la fase S se realiza la duplicación de los cromosomas en el núcleo, por replicación de las moléculas de ADN. Las dos moléculas hijas de cada cromosoma son identicas entre sí, por lo que se denominan cromátidas hermanas.
Durante esta fase se inicia la condensación de los cromatidas hermanas hasta formar los cromosomas mitóticos. Cada cromátida tiene forma de bastocillo y permanecen unidas entre si a través de una región denominada centromero.
La mitosis es la fase en la cual los núcleos de las células se dividen y se reparten las cromátidas hermanas. Es un proceso continuo en el que se distinguen cinco etapas: Profase, prometafase, metafase, anafase y telofase.
12.1 Profase
A lo largo de esta fase finaliza la condensación de las cromátidas hermanas iniciada en la fase G2. Los cromosomas mitóticos se van haciendo más cortos y gruesos, haciéndose visibles al microscopio óptico.
El complejo centriolar (formado por dos centriolos y fibras pericentriolares), se duplica formando los ásteres que comienzan a alejarse a polos opuestos del núcleo, al tiempo que se forman nuevas fibras de microtúbulos polares que constituyen el huso mitótico. Las células animales, que poseen centriolos y por tanto forman aster realizan una mitosis astral, para diferenciarla de la mitosis anastral que realizan las células vegetales que carecen de centriolos. En este caso las fibras del huso se forman en una región del citoplasma cercana al núcleo y libre de orgánulos que se conoce como zona clara.


La ruptura de la envoltura nuclear señala el comienzo de esta fase, quedando los cromosomas libres.
Se forman los microtúbulos cinetocóricos, a partir de los cinetocoros de los cromosomas. Estos microtúbulos son perpendiculares al eje del cromosoma.
Al final de esta fase los cromosomas comienzan a orientarse respecto a los polos, de modo que cada cinetocoro se encuentra frente a un de los polos.

Los cromosomas se disponen con sus cinetocoros en un mismo plano ecuatorial equidistantes de los dos polos, formando la llamada placa metafásica. Los microtúbulos cinetocóricos se alargan haca los polos.


12.4 Anafase
Las cromátidas hermanas de cada cromosoma se separan y se convierten en cromosomas independientes que son desplazados, a la misma velocidad y simultáneamente, hacia el polo que miran sus cinetocoros.
Esto es posible por un acortamiento de los microtúbulos cinetocóricos y un alargamiento de los microtubulos polares que alargan el huso y alejan ambos polos.


12.5 Telofase
Desaparecen las microtúbulos cinetocóricos y fragmentos de retículo endoplasmático se unen a los cromosomas, formando un esbozo de la envoltura nuclear.
Los microtúbulos polares se agrupan en haces y se alejan de los polos.
Al final de esta fase se completa la envoltura nuclear, los cromosomas comienzan a descondensarse y los microtúbulos polares se encuentran agrupados formando un haz único.


12.6 Citocinesis
La mitosis, en sentido estricto, es la división del núcleo celular pero también debe producirse la división del citoplasma y la separación en dos, o más, células hijas: esto es la citocinesis. Debido a la existencia de la pared celular el proceso es diferente en las células animales y las células vegetales:

12.6.1 Citocinesis en las células animales
Al final de la anafase, la membrana citoplasmática se invagina, gracias al anillo contráctil de fibras de actina dispuesto alrededor de la célula, iniciando así la formación del surco de división.
A lo largo de la telofase el surco crece y se comprime alrededor del haz de microtúbulos polares.
Al final de la telofase las células hijas solo están unidas por un puente citoplasmático donde se encuentra el haz de microtúbulos.
Por último, el puente citoplasmático se rompe, las células se separan y una de ellas arrastra los restos del haz de microtúbulos que finalmente serán degradados.
12.6.2 Citocinesis en las células vegetales
Durante la telofase, vesículas provenientes del aparato de Golgi confluyen en la región media de la célula y se fusionan formando el fragmoplasto.
Al final de la telofase todas las vesículas se han fusionado formando las membranas plasmáticas de las dos células hijas. A partir del contenido de la vesículas se forma la pared celular (lámina media). Los puentes o poros citoplasmáticos persisten entre las dos células hijas y constituyen los plasmodesmos.

Esta división nuclear es especial y su función es formar los gametos (imprescindibles para llevar a cabo la reproducción sexual). Los gametos son células haploides (n) con una sola copia de cada cromosoma mientras el resto de nuestras células son diploides (2n) con dos copias de cada cromosoma. Por eso a la meiosis muchas veces se le denomina división reduccional.
La meiosis va a consistir en dos divisiones consecutivas: una primera división meiótica en la que van a separarse los pares de cromosomas y una segunda división meiótica donde van a separarse las dos cromátidas hermanas de cada cromosoma (de forma similar a como ocurría en la mitosis).
13.1 Primera división meiótica
La dividimos en cuatro fases: profase I, metafase I, anafase I y telofase I (el I se indica siempre en numeración romana y no latina).
13.1.1 Profase I
Es la fase clave de la meiosis y la más larga y compleja. Durante todo el proceso la envoltura nuclear permanece intacta, al tiempo que se desintegra el nucleolo y se forma el huso mitótico. Se distinguen cinco fases:
a) Leptoteno; los cromosomas se hacen visibles por condensación. Sin embargo no se distinguen las cromatidas hermanas pues permanecen estrechamente unidas. Los cromosomas forman bucles de manera que sus extremos se unen a la cara interna de la envoltura nuclear a través de la placa de unión.


b) Zigoteno; los cromosomas homólogos se aparean punto por punto, de forma intima, en toda su longitud. Este emparejamiento gen a gen de los cromosomas homólogos se denomina sinapsis. Cada par cromosómico resultante de la sinapsis se denomina cromosoma bivalente o tétrada.
El apareamiento preciso de los homólogos queda estabilizado y mantenido hasta el final de la siguiente fase gracias a la formación de una estructura proteica en forma de cinta denominada complejo sinaptonémico.
La sinapsis se produce entre todos los cromosomas homólogos excepto entre los cromosomas X e Y, que solo se aparean parcialmente.

c) Paquiteno; los cromosomas se encogen haciéndose más gruesos. Durante esta fase se produce el intercambio de fragmentos cromatídicos entre cromátidas no hermanas de los cromosomas homólogos, mediante un procesos complejo conocido como entrecruzamiento y que es la causa de la recombinación genética. Este proceso es muy importante pues aumenta enormemente la variabilidad genética.
Las cromátidas hermanas ya son claramente visibles, de manera que cada cromosoma bivalente se observa claramente como una tétrada.

d) Diploteno; desaparece el complejo sinaptonémico lo que provoca la separación de los cromosomas homólogos. La separación no es total pues las cromátidas no hermanas permanecen unidas por zonas denominadas quiasmas, que son los puntos por donde se produjo el entrecruzamiento.



e) Diacinesis; la condensación de las tétradas es máxima. Las cromátidas no hermanas permanecen unidas por los quiasmas que se han desplazado a los extremos de los cromosomas homólogos. Al final de esta fase comienza la desintegración de la envoltura nuclear y del nucleolo, al tiempo que se duplica el nucleosoma, se forma el aster y se inicia la formación del huso con microtúbulos polares.

13.1.2 Metafase I
Se forman los microtúbulos cinetocóricos, pero a diferencia de la mitosis, los dos cinetocoros de un mismo cromosoma homólogo están orientados hacia el mismo polo, mientras que los cinotocoros del homólogo se orientan hacia el polo opuesto.
Esto ocurre porque se van a separar los cromosomas homólogos y no las cromátidas. Es muy importante no confundirlo (y el dibujo) con la metafase de la mitosis.

13.1.3 Anafase I
Los quiasmas se rompen y, al acortarse los microtúbulos cinetocóricos, se produce la separación de los cromosomas homólogos, que migran hacia polos opuestos.

13.1.4 Telofase I
Desaparece el huso y se forma la envoltura nuclear alrededor de los dos núcleos hijos. Los cromosomas se sufren una ligera descondensación y la célula se divide en dos (citocinesis).

Como resultado de esta primera división meiótica se producen dos células hijas que tienen cada una de ellas un juego completo de cromosomas homólogos procedentes de la célula madre (haploides). Estos cromosomas homólogos ya no son completamente paternos o maternos pues debido al entrecruzamiento alternan algunos segmentos paternos y maternos al azar. De esta forma la recombinación genética hace que todos los gametos sean genéticamente diferentes.

13.2 Segunda división meiótica
Una vez terminada la primera división meiótica se produce un breve periodo de interfase (sin síntesis de ADN) en el que los cromosomas se descondensan un poco pero enseguida vuelven a condensarse y se desencadena la segunda división meiótica (muy similar a la mitosis que ya hemos estudiado).
Vamos a distinguir cuatro fases principales: profase II, metafase II, anafase II y telofase II; seguidas por la división del citoplasma o citocinesis.
a) Profase II; es una fase muy breve pues los cromosomas ya estaban muy condensados. Se rompe la envoltura nuclear y se forma el huso mitótico.

b) Metafase II; los cromosomas se alinean en la placa metafásica disponiéndose de tal manera que cada cromátida hermana está enfocada a uno de los polos.

c) Anafase II; se separan las cromátidas hermanas y comienzan a migrar hacia los polos.

d) Telofase II; se forman las envolturas nucleares alrededor de los cuatro núcleos haploides

Al final se produce la citocinesis ocasionando las cuatro células hijas haploides correspondientes.

14. Principales diferencias entre la mitosis y la meiosis




No hay comentarios:
Publicar un comentario